Download
1 / 39
390 likes | 417 Views
Explore the intricate world of DNA, nucleosomes, and chromosomal organization through the lens of groundbreaking research. Learn about the nucleosome positioning code and its impact on gene regulation. Discover how DNA sequences influence nucleosome formation and understand the role of nucleosomes in gene expression.

E N D
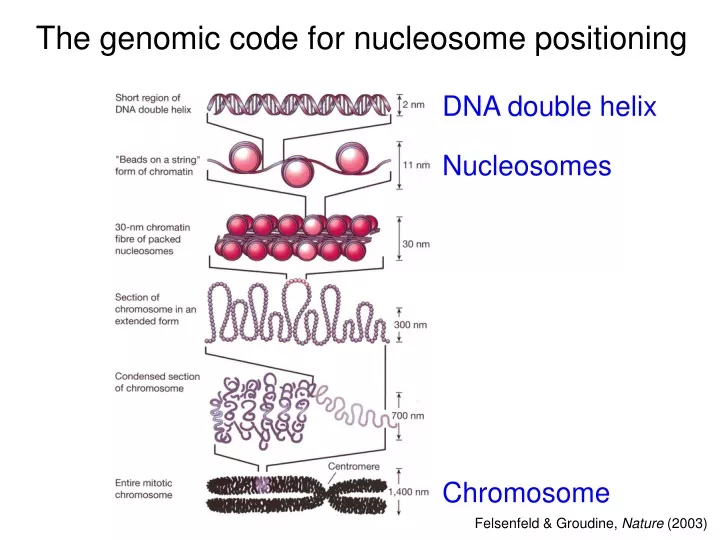
DNA double helix Nucleosomes Chromosome The genomic code for nucleosome positioning Felsenfeld & Groudine, Nature (2003)
DNA in nucleosomes is extremely sharply bent Side view (Space filling representation) Top view (Ribbon representation) ~80 bp per superhelical turn Luger et al., Nature (1997)
The nucleosome positioning code Nucleosomes like forming on this DNA sequence; CCAGCACCACCTGTAACCAATACAATTTTAGAAGTACTTTCACTTTGTAACTGAGCTGTCATTTATATTGAATTTTCAAAAATTCTTACTTTTTTTTTGGATGGACGCAA Nucleosomes dislike forming on this DNA sequence; ACTCTCCTCCGTGCGTCCTCGTCTTCACCGGTCGCGTTCCTGAAACGCAGATGTGCCTCCAATCGCACTGCTCCGAACAATAAAGATTCTACAATACTAGCTTTTATGGT
The nucleosome positioning code CCAAT Nucleosomes like forming on this DNA sequence; CCAGCACCACCTGTAACCAATACAATTTTAGAAGTACTTTCACTTTGTAACTGAGCTGTCATTTATATTGAATTTTCAAAAATTCTTACTTTTTTTTTGGATGGACGCAA Access of proteins to target site is hindered Nucleosomes dislike forming on this DNA sequence; ACTCTCCTCCGTGCGTCCTCGTCTTCACCGGTCGCGTTCCTGAAACGCAGATGTGCCTCCAATCGCACTGCTCCGAACAATAAAGATTCTACAATACTAGCTTTTATGGT
The nucleosome positioning code CCAAT Nucleosomes like forming on this DNA sequence; CCAGCACCACCTGTAACCAATACAATTTTAGAAGTACTTTCACTTTGTAACTGAGCTGTCATTTATATTGAATTTTCAAAAATTCTTACTTTTTTTTTGGATGGACGCAA Access of proteins to target site is hindered CCAAT Nucleosomes dislike forming on this DNA sequence; ACTCTCCTCCGTGCGTCCTCGTCTTCACCGGTCGCGTTCCTGAAACGCAGATGTGCCTCCAATCGCACTGCTCCGAACAATAAAGATTCTACAATACTAGCTTTTATGGT Easy access of proteins to target site in this region
Deciphering the nucleosome positioning code • In vitro selection of nucleosome-favoring DNAs • Isolation of natural nucleosome DNAs
Physical selection for DNAs that attract nucleosomes Random sequence DNA synthesis (1 each of 5 x 1012 different DNA sequences) Make many copies by PCR Equilibrium selection of highest affinity 10% Extract DNA Clone, sequence, analyze individuals Lowary & Widom, 1998
Summary • Differing DNA sequences exhibit a > 5,000-fold range of affinities for nucleosome formation Lowary & Widom, 1998 Thåström et al., 1999 Widom, 2001 Thåström et al., 2004
AATTTA GC AATTTA GC AATTTA GC GC AATTTA AATTTA GC GC AATTTA AATTTA GC DNA sequence motifs that stabilize nucleosomes and facilitate spontaneous sharp looping Thåström et al., 2004 Cloutier & Widom 2004 Segal et al., 2006
Isolation of natural nucleosome DNAs Digest unwrapped DNA Extract protected DNA Clone, sequence, analyze individuals
Fraction (AA/TT/TA) Position on nucleosome (bp) The nucleosome signature in living yeast cells • ~10 bp periodicity of AA/TT/TA • Same period for GC, out of phase with AA/TT/TA • Same signals from the in vitro nucleosome selection • NO signal from randomly chosen genomic regions Segal et al., 2006
Center alignment Location mixture model alignment Two alignments of nucleosome DNAs Wang & Widom, 2005
The nucleosome signature is common to yeast and chickens Chicken + Yeast merge Chicken (in vivo) Yeast (in vivo) Segal et al., 2006
Mouse (in vitro) Random DNA (in vitro) Yeast (in vitro) Chicken (in vivo) Yeast (in vivo) The nucleosome signature in vitro and in vivo Segal et al., 2006
In vitro experimental validationof histone-DNA interaction model • Adding key motifs increases nucleosome affinity • Deleting motifs or disrupting their spacing decreases affinity Segal et al., 2006
Summary Differing DNA sequences exhibit a > 5,000-fold range of affinities for nucleosome formation We have a predictive understanding of the DNA sequence motifs that are responsible Sequences matching these motifs are abundant in eukaryotic genomes, and are occupied by nucleosomes in vivo
Log likelihood Genomic Location (bp) Placing nucleosomes on the genome A free energy landscape, not just scores and a threshold !! • Nucleosomes occupy 147 bp and exclude 157 bp Segal et al., 2006
Equilibrium configurations of nucleosomeson the genome • One of very many possible configurations P(S) PB(S) P(S) PB(S) P(S) PB(S) P(S) PB(S) Chemical potential – apparent concentration Probability of placing a nucleosome starting at each allowed basepair i of S Probability of any nucleosome covering position i ( average occupancy) Locations i with high probability for starting a nucleosome ( stable nucleosomes) Segal et al., 2006
Binding sites for Gal4 activator protein GAL10 GAL1 147 bp Reading the nucleosome code and predicting the in vivo locations of nucleosomes Segal et al., 2006
Summary Differing DNA sequences exhibit a > 5,000-fold range of affinities for nucleosome formation We have a predictive understanding of the DNA sequence motifs that are responsible Sequences matching these motifs are abundant in eukaryotic genomes, and are occupied by nucleosomes in vivo A model based only on these DNA sequence motifs and nucleosome-nucleosome exclusion explains ~50% of in vivo nucleosome positions
Model Permuted Semi-stable nucleosomes Semi-stable nucleosomes Stable nucleosome TATA Box Distinctive nucleosome occupancy adjacent to TATA elements at yeast promoters Segal et al., 2006
Predicted nucleosome organization near 5’ ends of genes – comparison to experiment Segal et al., 2006 Fondufe-Mittendorf, Segal, & JW
Summary Differing DNA sequences exhibit a > 5,000-fold range of affinities for nucleosome formation We have a predictive understanding of the DNA sequence motifs that are responsible Sequences matching these motifs are abundant in eukaryotic genomes, and are occupied by nucleosomes in vivo A model based only on these DNA sequence motifs and nucleosome-nucleosome exclusion explains ~50% of in vivo nucleosome positions These intrinsically encoded nucleosome positions are correlated with, and may facilitate, essential aspects of chromosome structure and function
AATTTA GC AATTTA GC AATTTA GC GC AATTTA AATTTA GC GC AATTTA AATTTA GC An elastic energy model for the sequence-dependent cost of DNA wrapping Morozov, Fortney, Widom, & Siggia
DNA in nucleosomes is extremely sharply bent Side view (Space filling representation) Top view (Ribbon representation) ~80 bp per superhelical turn Luger et al., Nature (1997)
AATTTA GC AATTTA GC AATTTA GC GC AATTTA AATTTA GC GC AATTTA AATTTA GC An elastic energy model for the sequence-dependent cost of DNA wrapping Morozov, Fortney, Widom, & Siggia
E0 = Energy at equilibrium conformation for step fij = elastic constants impeding deformation; calculated from dispersion of parameters in X-ray crystal structures, assuming harmonic potential i = i – i0, = fluctuation of step parameter from equilibrium Elastic energy of dinucleotide step • Knowledge-based harmonic potential Olson et al., (1998)
Elastic energy model for nucleosomal DNA E = Eelastic + Edeviation from superhelix Crystal structure Ideal superhelix Morozov, Fortney, Widom, & Siggia
A genomic code for higher order chromatin structure? 30 nm fiber Felsenfeld & Groudine, 2003
Regular 3-d superstructures favor ~10 bp quantized linker DNA lengths Widom, 1992
Pairwise distances histogram(stable nucleosomes) Auto-correlations(average occupancy) Stable nucleosomes (model) Frequency Stable nucleosomes (permuted) Correlation Center-center distance (bp) Correlation offset (bp) Stable nucleosomes come in correlated groups Segal et al., 2006
Fourier transforms in extended regions Averaged for extended regions starting i = 11,…20 bp beyond end of mapped nuclesome: Period with max amplitude = 10.2 bp Phase offset at max period = 5 bp Wang, Fondufe-Mittendorf, & Widom
Digest linker DNA Isolate dinucleosomes Clone & sequence Biochemical isolation of dinucleosomes Yao et al., 1990; Fondufe-Mittendorf, Wang, & Widom
L’ N L N L’’ Linker lengths in purified dinucleosomes Predict locations of the two nucleosomes • Duration hidden Markov model: L’, N, L, N, L’’ L: Linker N: Nucleosome L’, L’’: Partial linkers Wang, Fondufe-Mittendorf, & Widom
The genomic code for nucleosome positioning DNA Nucleosomes 30 nm fiber Felsenfeld & Groudine, 2003
Multiplexing Layering two or more signals on top of each other without cross-interference • Multiple phone conversations in a single wire • or optical fiber • Stereo broadcast on an FM channel • Text message hidden in a picture, in a spy novel
How is multiplexing accomplished? • Nucleosomes not evolved for highest affinity; many ways to have suboptimal affinity over 147 bp length • Protein coding sequences and gene regulatory sequences are degenerate • A remarkable feature of DNA mechanics
+ Nucleosomes – nucleosomes Evolution of the nucleosome positioning code Sandman & Reeve, Curr. Op. Microbiol. 2006
Acknowledgements The genomic code for nucleosome positioning Northwestern University Yvonne Fondufe-Mittendorf Irene Moore Lingyi Chen Karissa Fortney Annchristine Thåström Timothy Cloutier Peggy Lowary Jiping Wang (NU Statistics Dept.) Weizmann Institute Eran Segal Yair Field Rockefeller University Eric Siggia Alexandre Morozov UCLA Robijn Bruinsma Joe Rudnick David Schwab