Download

1 / 18
280 likes | 929 Views
Chapter 7 Hemoglobin: Portrait of a Protein in Action. Heme = Fe 2+ (ferrous) + Protoporphyrin Protoporphyrin = 4 pyrrole rings + 4 methine bridges + 4 methyl groups + 2 vinyl groups + 2 propionate side chains

E N D
Chapter 7 Hemoglobin: Portrait of a Protein in Action
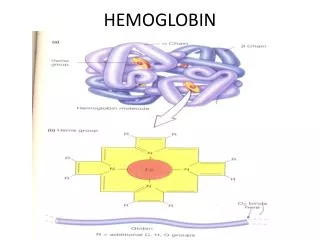
Heme = Fe2+ (ferrous) + Protoporphyrin • Protoporphyrin = 4 pyrrole rings + 4 methine bridges + • 4 methyl groups + 2 vinyl groups + • 2 propionate side chains • 5th coordination site occupied by the imidazole ring of proximal histidine of myoglobin (or hemoglobin) • 6th coordination site occupied by oxygen Heme Plane Shift upon O2 Binding Heme Allows Myoglobin and Hemoglobin to Bind Oxygen Max Perutz &John Kendrew First protein crystal structure in 1950s.1MBD.pdb by rearrangement of Fe electron
Maintenance of Heme Functionality Ferrous Oxymyoglobin [Fe2+/O2] Ferrous Deoxymyoglobin [Fe2+] + O2 O toxicity Ferric Oxymyoglobin [Fe3+/O2-] Metmyoglobin [Fe3+] + O2- X (unable to bind O2) • Oxygen must leave as dioxygen rather than superoxide. Why? • Reversible oxygen binding and storage • The hydrogen bonding between the distal histidine and oxygen stabilizes the ferric (Fe3+) form of oxymyoglobin and does not allow superoxide release. • Oxygen can be released only from the ferrous (Fe2+) form of oxymyoglobin.
Tetrameric Structure of Hemoglobin Max Perutz, horse heart 1A3N.pdb • Hemoglobin a Chain vs. Myoglobin : 25% Identity • Hemoglobin b Chain vs. Myoglobin : 24% Identity • Hemoglobin subunits and myoglobins share an evolutionarily conserved structural pattern called “globin fold”.
Mb : Hyperbolic P50 = 2 torr Hb : Sigmoidal P50 = 26 torr Oxygen Binding Properties of Myoglobin and Hemoglobin • 2,3-bisphosphoglycerate in red blood cell significantly lowers oxygen binding affinity of hemoglobin. • 4 independent oxygen bindings ? • Cooperative bindings ?
Loading & Unloading of Oxygen by Hemoglobin X10 than myoglobin, X1.7 than noncooperative protein • O2 Uptake by Hemoglobin in Lung (pO2 ~100 torr; 98% occupancy) • O2 Release in Typical Peripheral Tissues (pO2 ~20 torr; 32% occupancy; 66% Release) • O2 Unloading in Resting Muscle (pO2 ~40 torr; 77% occupancy; 21% Release) • O2 Unloading in Exercising Muscle (pO2 ~20 torr; 32% occupancy; 67% Release)
O2 Binding Changes the Quaternary Structure of Hemoglobin • 15 degree rotation of a1b1 dimer and a2b2 dimer upon oxygen binding • Overall structure of dimer themselves is relatively unchanged. • The interface between a1b1 dimer and a2b2 dimer is significantly changed. • Deoxyhemoglobin : T (for Tense) State; Stronger Inter-Subunit Interactions • Oxyhemoglobin : R (for Relaxed) State; Weaker Inter-Subunit Interactions • In R state, oxygen binding sites are free of strain and show higher affinity
Hemoglobin Cooperativity (1) Concerted Model (MWC Model) • Oxygen Binding Affinity : T state < R state • Oxygen binding shifts the quaternary structure of hemoglobin from T to R. • Oxyhemoglobin favors R state, whereas deoxyhemoglobin is in T state. • This model postulates that all 4 subunits are in the same conformation.
Oxygen Binding Affinity : square state ( ) < quarter circle state ( ) • Oxygen binding modulates the tertiary structure of the neighboring subunits. • This model postulates that the individual subunits can have different conformations. Hemoglobin Cooperativity (2) Sequential Model
Hemoglobin Cooperativity (3) The Truth ? • Neither the concerted model nor the sequential model is fully accurate. • Oxygen Binding Affinity Hemoglobin-[O2]0 (T) : Hemoglobin-[O2]3 (R) = 1 : 20 • (Concerted Model Works !!!) • Oxygen Binding Affinity Hemoglobin-[O2]0 (T) : Hemoglobin-[O2]1 (T) = 1 : 3 • (Sequential Model Works !!!)
Oxygen Binding to Heme Changes Interface Structures • Oxygen binding causes T to R quaternary structure changes. HOW ??? • Oxygen binding induces heme plane movement. • Heme plane movement prompts conformational changes of hemoglobin subunit via proximal histidine residue, which is the 5th coordination site of the heme iron. • The C-terminal end of the individual subunit, which is located in the inter-subunit space, is relocated upon oxygen binding. • The structural transition at the iron in one subunit is directly transmitted to the other subunits changing the interface between the ab dimers. • Cooperative oxygen binding by hemoglobin
Specific Binding of 2,3-BPG to T-State Hemoglobin • The T state of hemoglobin is highly unstable. (themodynamically not favored…) • R T transition is essential for efficient oxygen unloading. • Thus, an additional mechanism is necessary to promote R T transition !!! • In red blood cell, [Hb] = [2,3,-BPG] = ~2 mM. • 2,3-BPG can bind to the central pocket located in the center of the tetramer. • 2,3-BPG binding pocket can be generated only in the T-state. • Thus, 2,3-BPG can stabilize the T form population.
Cargo Transport Efficiency of Hemoglobin vs. 2,3-BPG • [2,3-BPG]LOW R > T and [2,3-BPG]HIGH R < T (Allosteric Regulation) • O2 unloading efficacy 2,3-BPG(-) : 2,3-BPG(+) = 8% : 66% • Adult Hemoglobin [ab]2 vs. Fetal Hemoglobin [ag]2 • In g chain, serine residue has replaced 143 Histidine residue of b chain, which provides a positive charge critical for optimum 2,3-BPG binding. • Thus, 2,3-BPG binding is much less efficient for fetal hemoglobin. • Oxygen affinity : fetal hemoglobin > maternal hemoglobin.
The Bohr Effect (1) : Hydrogen Ion (pH) • Rapidly metabolizing tissues such as contracting muscle generate large amounts of hydrogen ions. • Decrease of pH from 7.4 to 7.2 increases oxygen unloading efficiency by 11%. • Salt bridges involving Lys40 of a2, His146 of b1 and Asp94 of b1 stabilize the T form of hemoglobin. • Protonation on His146 of b1 enhances salt bridge formation with Asp94 of b1.
The Bohr Effect (2) : Carbon Dioxide (1) Constant pH! • Actively metabolizing tissues release large amounts of CO2 resulting in pH decrease. • Carbonic Anhydrase : Carbon Dioxide Carbonic Acid (pKa 3.5) Bicarbonate + H+ • 40 torr CO2increases oxygen unloading efficiency by 11% at pH 7.2. CO2 decrease the affinity of hemoglobin for oxygen beyond the effect due to decrease in pH
The Bohr Effect (3) : Carbon Dioxide (2) • CO2 can react with the N-terminal amino group of the hemoglobin subunits and yield carbamate groups, which produce additional negative charges. • The amino termini reside in the interface of ab dimers. • These CO2-induced negative charges in the N-termini of hemoglobin subunits consequently promote salt bridges formation stabilizing the T form.
Carbon Dioxide Deportation • Carbamate yielding reactions contribute to process 14% of the total CO2 released from the peripheries. • The majority of the CO2 released from the peripheries is uptaken by the red blood cells and processed to HCO3-, and eventually exhaled in lung as CO2 via reverse conversion reactions (i.e. HCO3- CO2).
Sickle-Shaped RBC Hemoglobin Fibers Deoxyhemoglobin S Sickle Cell Anemia • In Sickle-Cell Anemia, Glu6 of b chain is mutated to Val6 (HbS substitution). • The deoxy HbS b chains can establish aberrant hydrophobic interactions involving Val6 of one b chain and Phe85 and Val88 of another b chain (i.e. between different hemoglobin tetramers). Deoxyhemoglobin polymerization Formation of hemoglobin fibers (i.e. insoluble aggregates of hemoglobin) • The HbS substitution does not alter the properties of oxyhemoglobin because the hydrophobic interaction between Val6 and Phe85/Val88 is not allowed in R state.