Download

1 / 22
220 likes | 439 Views
Modelling of pyruvate decarboxylases from ethanol producing bacteria. Anjala Shrestha 1 , Srisuda Dhamwichukorn 1 , Ekachai Jenwitheesuk 2* 1 Joint Graduate School of Energy and Environment, KMUTT, Thailand

E N D
Modelling of pyruvate decarboxylases from ethanol producing bacteria Anjala Shrestha1, Srisuda Dhamwichukorn1, Ekachai Jenwitheesuk2* 1 Joint Graduate School of Energy and Environment, KMUTT, Thailand 2National Center for Genetic Engineering and Biotechnology, 113 Thailand Science Park, Thailand Shrestha et al.,
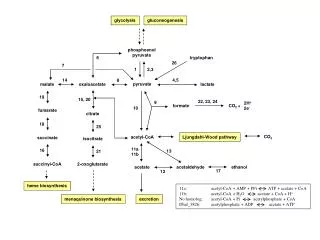
IntroductionPyruvate decarboxylase (PDC, EC 4.1.1.1) • Common in plants and fungi but very rare in prokaryotes and absent in animals (Konig,1998 ). • Non-oxidative Decarboxylation of 2-keto acid to aldehydes • Carboligation with aldehyde to form chiral 2-hydroxyketones (Pohl, 2004) • Carboligation with benzaldehyde to form R-phenylacetylcarbinol (R-PAC) Shrestha et al.,
Catalysis steps of PDC • Thiamin diphosphate(ThDP) and Mg2+ ions as cofactors. Shrestha et al.,
PDC possessing microorganisms Gram-negative- Zymomonas mobilis (ZmPDC) -Zymobacter palmae (ZpPDC) Gram-positive -Sarcina ventriculi (SvPDC) Possess Pyruvate decarboxylase and alcohol dehydrogenase (adh, EC 1.1.1.1) Z. mobilisATCC29291PDB ID: 1zpd (Dobritzsch, 1998) Shrestha et al.,
Objective of studyTo generate 3D structures of the ZpPDC and SvPDC using homology modelling techniqueEnzyme-substrate interactionsSubunit-subunit interfaces that might be related to the different biochemical characteristics. Shrestha et al.,
Methodology Predicted 3D-structures Model assessment Discrete Optimized Protein Energy Residue-specific All-atoms Conditional Probability Discriminatory Function Modeller 9v3 (Sali, 1993) Shrestha et al.,
Energy minimization by the NAMD program without water molecules (Phillips, 2005) • Protein interface analysis by Protein-Protein interaction server (Reynolds, 2009) • PROCHECK version 3.5.4 (Morris,1992) • Docking of cofactors by superimposition with the template by DeepView (SWISS-PdbViewer, v. 3.7) • Mol_Volume version 1 (Kalé, 1999), R_PROBE-2.0 Å. Shrestha et al.,
Protein-ligand docking by Autodock 4 (Morris, 1998) Autodock graphical user interface (Autodock tools 1.4.6) Protein, Ligand structures Grid box (40 points on each side with distance of 0.375 Å) AutoGrid version 4 Affinity Maps for each atom types of ligands Ki= exp [(∆G X 1000)/ (Rcal X TK) ∆G= Intermolecular energy + Internal energy of ligand Rcal=1.98719 cal K-1 mol-1 TK=298.15 K, Kd=1/Ki Autodock version4 Shrestha et al.,
Results and DiscussionsHomology models assessment Cα : ZpPDC and ZmPDC 2.2.7 Å (Picture A) SvPDC and EcIPDC 2.93 Å (Picture B) Shrestha et al.,
Ramachandran Plot ZpPDCSvPDC Residue in most favoured regions 879 (90.4%) 882 (88.6%) Residue in additional allowed regions 86 (8.8%) 103 (10.3) Residues in generously allowed regions 5 (0.5%) 7 (0.7%) Residues in disallowed regions 2 (0.2%) 4 (0.4%) Overall PROCHECK (-1.0 to - 0.05) -0.32 -0.33 Shrestha et al.,
Secondary Structures • α/β topology common to all thiamine dependent enzymes. ZpPDC-PYR ZpPDC-R ZpPDC-PP Shrestha et al.,
Interface Analysis of Models • H-bonds • SvPDC model (30) • ZpPDC model (44) Interface areas • ZpPDC= 3587 Å2 • SvPDC = 3284 Å2 R2=0.75 Conformational changes upon substrate binding Shrestha et al.,
Binding modes of the cofactors Residues within 4 Å of ThDP X-ray crystal str. of ZmPDC Homology model of ZpPDC Homology model of SvPDC Blue: Acidic; Yellow: Polar Red: Basic; Grey: Hydrophobic Shrestha et al.,
Substrate binding sites of the homology models Asp26*, His113*, His114*, Tyr290, Thr388 and Glu473 in ZmPDC Ala in EcIPDC & ScPDC Tyr384 in SvPDC SvPDC: Thr288 Conversion of Thr288 to ZmPDC analogue would decrease active site volume and subsequently decrease Km value for pyruvate. Shrestha et al.,
Pyruvamide interacts with ThDP and side chains of Asp26, His113, Glu468 of ZpPDC, and Asp27, His114 of SvPDC. The distances between pyruvamide and these structures are within 4 Å . Shrestha et al.,
Substrate activation of homology models • SvPDC model suggests that conformational changes occurs upon substrate binding at regulatory site similar to ScPDC form-B (Lu, 2000). Shrestha et al.,
Affinity of ZpPDC and SvPDC towards other 2-keto acids Linear relationship between molecular volume of substrate and experimental Km value for ZmPDC Shrestha et al.,
The larger 2-keto acids bind with higher dissociation constant (Kd =1/Ki) than those of the smaller 2-keto acids to both homology models. Ile472 and Thr388 of ZmPDC Substrate binding preferences of the ZpPDC and SvPDC enzymes are similar to ZmPDC Shrestha et al.,
Conclusion • Homology models of PDCs of Z. palmae and S. ventriculi -- to explore the structural similarities and differences among bacterial PDCs at the atomic level. • Similar in cofactor binding modes, substrate binding residues, and active site volume. • Mechanism of allosteric activation shown by SvPDC is similar to ScPDC form-B which needs further experimental verification. • Preference of ZpPDC and SvPDC enzymes for aliphatic 2-ketoacids. Shrestha et al.,
Acknowledgment Joint Graduate School of Energy and Environment (JGSEE), King Mongkut’s University of Technology Thonburi, Bangkok, Thailand. Grateful to Philip Shaw, Sissades Tongsima, Pavita Tipsombatboon, Alisa Wilantho and Wanwimon Mokmak for their technical support and valuable comments. Shrestha et al.,
Thank you for your attention. Shrestha et al.,
ZmPDC 1 --MSYTVGTYLAERLVQIGLKHHFAVAGDYNLVLLDNLLLNKNMEQVYCCNELNCGFSAE 58 GYARAKGAAAAVVTYSVZpPDC 1 ---MYTVGMYLAERLAQIGLKHHFAVAGDYNLVLLDQLLLNKDMEQVYCCNELNCGFSAE 57 GYARARGAAAAIVTFSVSvPDC 1 --MKITIAEYLLKRLKEVNVEHMFGVPGDYNLGFLDYVEDSKDIEWVGSCNELNAGYAAD 58 GYARLRGFGVILTTYGVEcIPDC 1 MRTPYCVADYLLDRLTDCGADHLFGVPGDYNLQFLDHVIDSPDICWVGCANELNASYAAD 60 GYARCKGFAALLTTFGVScPDC 1 -MSEITLGKYLFERLKQVNVNTVFGLPGDFNLSLLDKIYEVEGMRWAGNANELNAAYAAD 59 GYARIKGMSCIITTFGV ▲ Pyrα4 Pyrβ4 Pyrα5 Pyrα6 Pyrβ5Pyrα7 ZmPDC 76 G-ALSAFDAIGGAYAENLPVILISGAPNNNDHAAGHVLHHALG 117 KTDYHYQLEMAKNITAAAEAIYTPEEAPAKIDHZpPDC 75 G-AISAMNAIGGAYAENLPVILISGSPNTNDYGTGHILHHTIG 116 TTDYNYQLEMVKHVTCARESIVSAEEAPAKIDHSvPDC 76 G-SLSAINATTGSFAENVPVLHISGVPSALVQQNRKLVHHSTA 117 RGEFDTFERMFREITEFQSIISEYN-AAEEIDREcIPDC 78 G-ELSAMNGIAGSYAEHVPVLHIVGAPGTAAQQRGELLHHTLG 119 DGEFRHFYHMSEPITVAQAVLTEQN-ACYEIDRScPDC 77 G-ELSALNGIAGSYAEHVGVLHVVGVPSISAQAKQLLLHHTLG 118 NGDFTVFHRMSANISETTAMITDIATAPAEIDRPyrβ6Pyrα8Rα1 Rβ1ZmPDC 151 VIKTALRE-KKPVYLEIACNIASMPCA 176 APGPASALFNDEASDEA-SLNAAVEETLKFIAXRDKVAVLVGSKLRAAGZpPDC 150 VIRTALRE-RKPAYLEIACNVAGAECV 175 RPGPINSLLRELEVDQT-SVTAAVDAAVEWLQDRQNVVMLVGSKLRAAASvPDC 150 VIESIYKY-QLPGYIELPVDIVSKEIE 175 IDEMK-PLNLTMRSNEK-TLEKFVNDVKEMVASSKGQHILADYEVLRAKEcIPDC 152 VLTTMLRE-RRPGYLMLPADVAKKAAT 177 PPVNA-LTHKQAHADSA-CLKAFRDAAENKLAMSKRTALLADFLVLRHGScPDC 152 CIRTTYVT-QRPVYLGLPANLVDLNVP 177 AKLLQTPIDMSLKPNDAESEKEVIDTILALVKDAKNPVILADACCSRHDRα2 Rβ2 Rβ3 Rα3Rα4 Rβ4 ZmPDC 225 AEEAAVKFADA 235 LGGAVATMAAA-KSFFPEENPHYIGTSWGEVSYPGVEKTMKEADAVIALAPVFNDYSTTG 294 WZpPDC 224 AEKQAVALADR 234 LGCAVTIMAAE-KGFFPEDHPNFRGLYWGEVSSEGAQELVENADAILCLAPVFNDYATVG 293 WSvPDC 223 AEKELEGFINE 233 AKIPVNTLSIG-KTAVSESNPYFAGLFSGETSSDLVKELCKASDIVLLFGVKFIDTTTAG 292 FEcIPDC 225 LKHALQKWVKE 235 VPMAHATMLMG-KGIFDERQAGFYGTYSGSASTGAVKEAIEGADTVLCVGTRFTDTLTAG 294 FScPDC 227 VKAETKKLIDL 237 TQFPAFVTPMG-KGSIDEQHPRYGGVYVGTLSKPEVKEAVESADLILSVGALLSDFNTGS 296 FRβ5 Rβ6 Rβ7 Rα5 Rα6 PPα1 ZmPDC 296 TDIPDPKKLVLAEPRSVVVNGIRFPSVHLKDYLTRLAQKVSKKTGALDFFKSLNAGELK 354 KAAPADPSAPLVNAEIAZpPDC 295 NSWPKGDNVMVMDTDRVTFAGQSFEGLSLSTFAAALAEKAPSRPATTQGTQAP----VL 349 GIEAAEPNAPLTNDEMTSvPDC 294 RYINKDVKMIEIGLTDCRIGETIYTGLYIKDVIKALTD------AKIKFHNDVKVEREA 346 VEKFVPTDAKLTQDRYFEcIPDC 296 THQLTPAQTIEVQPHAARVGDVWFTGIPMNQAIETLVEL-----CKQHVHAGLMSSSSG 349 AIPFPQPDGSLTQENFWScPDC 298 SYSYKTKNIVEFHSDHMKIRNATFPGVQMKFVLQKLLTTIAD--AAKGYKPVAVPARTP 354 ANAAVPASTPLKQEWMWPPβ1PPα2 PPβ2PPα3 PPβ3 /PPα4ZmPDC 372 RQVEALLTPNTTVIAETGDSWFNAQRMKLPNGARVEYEMQWGH 414 IGWSVPAAFGYAVGA----PERRNILMVGDGSFZpPDC 367 RQIQSLITSDTTLTAETGDSWFNASRMPIPGGARVELEMQWGH 409 IGWSVPSAFGNAVGS----PERRHIMMVGDGSFSvPDC 364 KQMEAFLKPNDVLVGETGTSYSGACNMRFPEGSSFVGQGSWMS 406 IGYATPAVLGTHLAD----KSRRNILLSGDGSFEcIPDC 367 RTLQTFIRPGDIILADQGTSAFGAIDLRLPADVNFIVQPLWGS 409 IGYTLAAAFGAQTAC----PNRRVIVLTGDGAAScPDC 372 NQLGNFLQEGDVVIAETGTSAFGINQTTFPNNTYGISQVLWGS 414 IGFTTGATLGAAFAAEEIDPKKRVILFIGDGSL ▲ -▲▲▲- PPα5 PPβ4 PPα6PPα7PPα8 PPβ5PPα9ZmPDC 444 QLTAQEVAQMVRLKLPVIIFLINNYGY 470 TIEVMIHDG--PYNNIKNWDYAGLMEVFNGNGGYDSGAGKGLKAKTGGEZpPDC 439 QLTAQEVAQMIRYEIPVIIFLINNRGY 465 VIEIAIHDG--PYNYIKNWNYAGLIDVFND----EDGHGLGLKASTGAESvPDC 436 QLTVQEVSTMIRQKLNTVLFVVNNDGY 462 TIERLIHGPEREYNHIQMWQYAELVKTLATE---RDIQPTCFKVTTEKEEcIPDC 439 QLTIQELGSMLRDKQHPIILVLNNEGY 465 TVERAIHGAEQRYNDIALWNWTHIPQALS-----LDPQSECWRVSEAEQScPDC 448 QLTVQEISTMIRWGLKPYLFVLNNDGY 474 TIEKLIHGPKAQYNEIQGWDHLSLLPTFG------AKDYETHRVATTGE -----------------------▲ ▲ ▲▲PPβ6 PPα10 .ZmPDC 518 LAEAIKVALAN 528 -TDGPTLIECFIGREDCTEELVKWGKRVAAANSRKPVNKLL 568ZpPDC 509 LEGAIKKALDN 519 -RRGPTLIECNIAQDDCTETLIAWGKRVAATNSRKPQA--- 556SvPDC 509 LAAAMEEINKG 519 -TEGIAFVEVVMDKMDAPKSLRQEASLFSSQNNY------- 552EcIPDC 510 LADVLEKVAH- 519 -HERLSLIEVMLPKADIPPLLGALTKALEACNNA------- 552ScPDC 518 WDKLTQDKSFN 528 DNSKIRMIEIMLPVFDAPQNLVEQAKLTAATNAKQ------ 563 Shrestha et al.,