Download

1 / 33
330 likes | 452 Views
Optimization of non-natural nucleotides for selective incorporation opposite damaged DNA. Diana Vineyard, Xuemei Zhang, Alison Donnelly, Irene Lee and Anthony J. Berdis. Org. Biomol. Chem. , 2007 , 5 , 3623–3630. Nucleic acid bases : Purines and Pyrimidines. Purines. Adenine. Guanine.

E N D
Optimization of non-natural nucleotides for selective incorporation oppositedamaged DNA Diana Vineyard,Xuemei Zhang,Alison Donnelly,Irene Leeand Anthony J. Berdis Org. Biomol. Chem., 2007, 5, 3623–3630
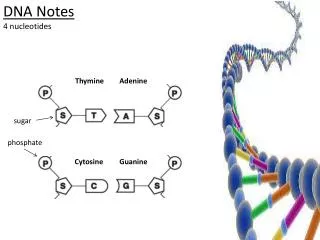
Nucleic acid bases :Purines and Pyrimidines Purines Adenine Guanine Pyrimidines Uracil (RNA) Cytocine Thymine (DNA)
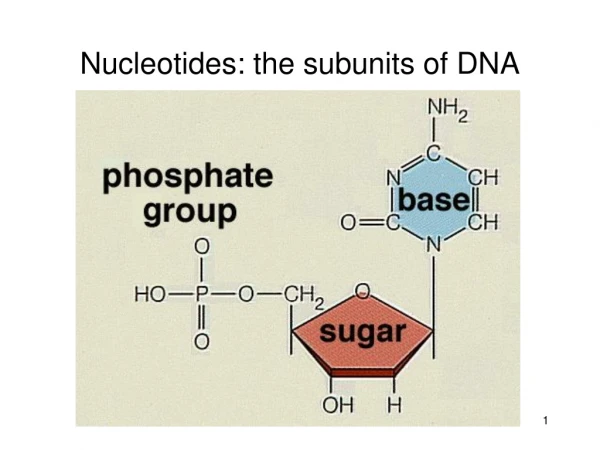
Structure of DNA : Bases linked to Sugar-Phosphate backbone Phosphodiester linkage
Structure of DNA : Double Helix Phosphates and Sugars: Hydrophilic Bases: Hydrophobic H – Bonding between Base pairs
Base Stacking Interactions • Hydrophobic stacking interactions in which two of more bases are positioned with the planes of their rings parallel • Also involve combination of van der Waals an dipole-dipole interactions between bases • Help to minimize contact with water and are very important in stabilizing 3D structure of DNA
DNA damages Oxidative damage UV induced damage: Thymine dimers Hogg, M.; Wallace, S. S.; Doublie, S. Current Opinion in Structural Biology 2005, 15, 86–93
DNA damages: Can be mutagenic if bypassed by DNA polymerase Normal C-G pair 8-oxoG –dC (dexoycytosine 3’-monophosphate) Mutagenic ! 8-oxoG –dA (dexoyadenosine 3’-monophosphate) Hogg, M.; Wallace, S. S.; Doublie, S. Current Opinion in Structural Biology, 2005, 15, 86–93
Abasic Sites • Most freaquent DNA lesion encountered by cells (~10,000 per human cell per day) • Produced by hydroxyl radical attack on the sugar residue releasing the free base or spontaneous depurination Hogg, M.; Wallace, S. S.; Doublie, S. Current Opinion in Structural Biology, 2005, 15, 86–93
DNA Polymerases • Belong to different families • Polymerase domain is composed of three sub domains: fingers, palm and thumb • The exonuclease domain increases proof- reading activity by10-100 fold
RB69 DNA Polymerase gp43:a close relative of T4 DNA polymerase gp43
DNA Replication • Substrate : deoxynucleoside 5’-triphosphates or dNTP (dATP, dGTP,dTTP,dCTP) dATP
Chemistry of DNA replication Pyrophosphate ion
Can abasic sites be bypassed by DNAPolymerase? Primer extended by DNA polymerase Primer not extended RB69 gp43 exo- is able to extend a primer by incorporating dAMP, and to a lesser extent dGMP, opposite an abasic site. Hogg, M.; Wallace, S. S.; Doublie, S. The EMBO Journal, 2004, 23, 1483–1493
Goal: translesion DNA synthesis Translesion DNA synthesis: The ability of a DNA polymerase to incorporate opposite a DNA lesion DNA substrate (primer): X= abasic site - part I or A, C, G, T (templating bases) – part II DNA polymerase: T4 DNA polymerase (gp43 exo-)
Surface-ionization potentials for each nucleobase Red: electronegative Green: neutral Blue: electropostive
dNTP analogs • 5-cyanoindolyl-2’-deoxyriboside 5’-triphosphate (5-CyITP) • 5-ethylindolyl-2’-deoxyriboside 5’-triphosphate (5-EtITP) • 5-ethyleneindolyl-2’-deoxyriboside 5’-triphosphate (5-EyITP) • 5-methylindolyl-2’-deoxyriboside 5’-triphosphate (5-MeITP)
Parameters Kd:The first kinetic step represents binding of dXTP to the polymerase–DNA complex Kpol: conformational change Kchem: The final stage of the catalytic cycle is the phosphoryl transfer step that is required for elongation of the primer strand Kobs: observed product formation
What physical properties dictate the incorporation of unnatural nucleotides opposite abasic sites?
Part I:Enzymatic incorporation opposite an abasic site Rate constants (Kobs) for the incorporation of 5-CyITP opposite an abasic site on as a function of nucleotide concentration Rate constants (Kobs) vs. [5-CyITP] Hyperbolic; Fitted to Mechaelis-Menten kinetics
Part I: Comparison of 5-CyITP with 5-NITP: oberservations • 5-NITP: 5-nitroindolyl-2’-deoxyriboside 5’-triphosphate • Note: two analogs are similar in size, solvation energies, dipole moments, and presence of π-electron density • kpol of 29 s−1 for 5-CyITP is ∼4-fold slower than the kpol value of 126 s−1 for • Kd for 5-CyITP (58μM) is 3-fold higher than 18 μM for 5-NITP • 5-NITP more efficiently incorporated?
Comparison of 5-CyITP with 5-NITP: explanations for Kpol • The nitrogen atom of the nitro substituent group of 5-NITP possesses a partial positive charge and therefore can interact with O4 of Thymine adjacent to abasic site(dipole-dipole interactions favor base stacking) * *
Comparison of 5-EyITP with 5-CEITP: observations • 5-CEITP: 5-cyclohexylindolyl-2’-deoxyriboside 5’-triphosphate • 5-EyITP binds10-fold more weakly than 5-CEITP • The faster kpol value of 94 s−1 for 5-EyITP compared to 25 s−1 for 5-CEITP • 5-CEITP more efficiently incorporated?
Comparison of 5-EyITP with 5-CEITP: explanations • Reduced size and hydrophobicity of 5-EyITP limits its ability to adequately displace any water molecules (faster Kd) that may occupy the void of the abasic site as effectively as the larger 5-CEITP • The faster kpol value of 94 s−1 for 5-EyITP compared to 25 s−1 for 5-CEITPlikely reflects the ability of the smaller analog to facilitate the conformational change preceding phosphoryl transfer more effectively than the larger 5-CEITP analog
Part I: Conclusion Nucleotide incorporation opposite a non-templating DNA lesion can occur via enhanced base-stacking interactions caused by reductions in solvation energies and proper steric arrangements rather than through overall increases in πelectron surface area
Part I: Conclusion How about π-electron surface area?
Part I: Conclusion – how to design selectivity of incorporation Selectivity for incorporation opposite an abasic site can be modulated through simple alterations to the π-electron surface area of a non-natural nucleotide Data fit parabolic function well
Part II:Enzymatic insertion opposite templating nucleobases • All small non-natural nucleotides synthesized in this study are poorly incorporated opposite any templating nucleobase
Selectivity of incorporation with respect to of a non-natural nucleotideopposite a templating base Catalytic efficiency vs. π-electron surface area: Parabolic curve is non-existent
Part II : Conclusions • Challenging to rationally design a nucleotide for selective incorporation opposite a templating nucleobase • Difficult to find a model explaining kinetic behavior of these non-natural nucleotides incorporated opposite templating nucleobases • Nucleotide incorporation opposite templating DNA is influenced by these interrelated biophysical features • Abasic site appears to reduce the overall complexity of these interactions such that the catalytic efficiency for incorporation is primarily influenced by only the size and π-electron density of the incoming nucleotide
Significance and future work • Designing new base analogs to control DNA polymerase activity – applying synthetic organic chemistry • More accurately define factors that control incorporation opposite a templating bases – physical chemistry • Better understand mechanism of DNA repair – DNA lesions play key role in cancer development
Acknowlegement • Dr. Martin Case • All the Chem 258 students