Download

1 / 33
330 likes | 362 Views
Explore the process of converting extracellular signals into cellular responses and the key components involved, including signaling molecules, receptors, and intracellular proteins. Learn about G protein-coupled receptors and different pathways used in cell signaling.

E N D
Chap. 15 Signal Transduction & G Protein-coupled Receptors • Topics • Signal Trans.: From Extracellular Signal to Cellular Response • Cell-Surface Receptors & Signal Transduction Proteins • G Protein-coupled Receptors (GPCRs): Structure and Mechanism • GPCRs That Regulate Ion Channels • GPCRs That Regulate Adenylyl Cyclase • GPCRs That Regulate Cytosolic Calcium • Goals • Learn the general properties of signaling molecules (ligands), cell-surface receptors, & intracellular signal transduction components. • Learn the G protein cycle of reactions involved in GPCR signaling. • Learn the rhodopsin signal trans pathway used in vision. • Learn the epinephrine receptor signal trans pathway used for control of glycogen degradation. • Learn about the GPCR-stimulated IP3/DAG signaling pathway.
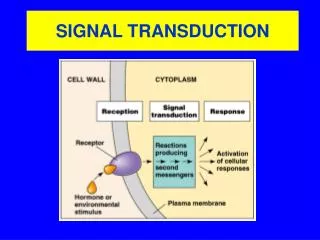
General Principles of Signal Transduction Signal transduction refers to the overall process of converting extracellular signals into intracellular responses (Fig. 15.1). Key players in signal transduction are signaling molecules, receptors, signal transduction proteins and second messengers, and effector proteins. Cells respond to signals by changing the activity of existing enzymes (fast) and/or the levels of expression of enzymes and cell components (slower) by gene regulation (Steps 7a & 7b). Receptors and signal transduction systems have evolved to detect and respond to hormones, growth factors, neurotransmitters, pheromones, oxygen, nutrients, light, touch, heat, etc. There are an enormous number of signal molecules and receptors in cells. In contrast, there are relatively few types of intracellular signal transduction systems.
General Principles of Signal Transduction In animals, signaling systems are classified based on the distance over which they act (Fig. 15.2). Endocrine signaling acts over long distances within the organism (e.g., insulin). Paracrine signaling acts over very short distances, for example between neighboring cells. Neurotransmitters and developmental signals typically act in this manner. In autocrine signaling, cells release ligands that bind to their own surface receptors, modulating activity. Many growth factors act in this manner. Finally, signaling systems involving plasma membrane-attached proteins act via direct cell-to-cell contact.
Signal Transduction Components: Receptors Cell surface receptors bind to their ligands (signaling molecules) via their extracellular domains (Fig. 15.3). In all cases, binding causes a conformational change in the receptor that leads to the transmission of an intracellular signal. Binding specificity and affinity are determined by the extent of molecular complementarity between the ligand and the receptor. A given receptor may exhibit specificity for a certain ligand or a group of closely related (structurally) ligands. A given ligand may bind to a number of different types of receptors, that exhibit different effector specificity (different cell responses). Further, two receptors that bind different ligands, may signal via the same intracellular signal transduction system, even within a single cell.
Signal Transduction Components: Kinases/Phosphatases Proteins that participate in intracellular signal transduction fall into two main classes--protein kinases/phosphatases and GTPase switch proteins. Kinases use ATP to phosphorylate amino acid side-chains in target proteins. Kinases typically are specific for tyrosine or serine/threonine sites. Phosphatases hydrolyze phosphates off of these residues. Kinases and phosphatases act together to switch the function of a target protein on or off (Fig. 15.4). There are about 600 kinases and 100 phosphatases encoded in the human genome. Activation of many cell-surface receptors leads directly or indirectly to changes in kinase or phosphatase activity. Note that some receptors are themselves kinases (e.g., the insulin receptor).
Model for Kinase-mediated Signal Trans. Fig. 15.5 illustrates a simple signal transduction pathway involving one kinase bound to a receptor and one predominant target protein. A number of signaling systems discussed in the course function via this general model.
Signal Trans. Components: GTPase Switches GTPase switch protein also play important roles in intracellular signal transduction (Fig. 15.6). GTPases are active when bound to GTP and inactive when bound to GDP. The timeframe of activation depends on the GTPase activity (the timer function) of these proteins. Proteins known as guanine nucleotide-exchange factors (GEFs) promote exchange of GTP for GDP and activate GTPases. Proteins known as GTPase-activating proteins (GAPs), stimulate the rate of GTP hydrolysis to GDP and inactivate GTPases. We will cover two classes of GTPase switch proteins--trimeric (large) G proteins, and monomeric (small) G proteins. Trimeric G proteins interact directly with receptors, whereas small G proteins interact with receptors via adaptor proteins and GEFs.
Signal Trans. Components: 2nd Messengers While there are a large number of extracellular receptor ligands ("first messengers"), there are relatively few small molecules used in intracellular signal transduction ("second messengers"). In fact, only 6 second messengers occur in animal cells. These are cAMP, cGMP, 1,2-diacylglycerol (DAG), and inositol 1,4,5-trisphosphate (IP3) (Fig. 15.8), and calcium and phosphoinositides (covered later). The functions of cAMP, cGMP, DAG, and IP3 are summarized in the figure. Second messengers are small molecules that diffuse rapidly through the cytoplasm to their protein targets. Another advantage of second messengers is that they facilitate amplification of an extracellular signal.
Signal Amplification in Signaling Pathways At each step of many signal transduction pathways, the number of activated participants in the pathway increases (Fig. 15.9). This is referred to as signal amplification, and hormone signaling pathways are often referred to as amplification cascades. For example, one epinephrine-activated GPCR activates 100s of Gas-GTP complexes, which in turn activate 100s of adenylyl cyclase molecules, that each produce hundreds of cAMP molecules, and so on. The overall amplification associated with epinephrine signaling is estimated to be ~108-fold.
Ligand Binding and Receptor Activation The reversible kinetic equation for ligand (L) binding to a receptor (R) is R + L RL The dissociation constant for this reaction is Kd = [R][L] / [RL]. When [L] ~ Kd, the receptor is ~50% saturated. When [L] = 10Kd, the receptor is ~90% saturated; at [L] = 0.1Kd, the receptor is ~10% saturated. Typically, the Kd for ligand binding is higher than the basal concentration of ligand. This is needed for cells to optimally respond to changing ligand concentration. Interestingly, the level of physiological response typically does not strictly parallel binding (Fig. 15.12). Namely, 50% of full response often occurs at only 10-20% receptor occupancy. The number of receptors in a cell is very important in setting the physiological response. A decrease in receptor number reduces the response, and vice versa. You are not responsible for the additional mathematical treatment of ligand-receptor binding covered in the text.
Ligand Agonists & Antagonists in Medicine Synthetic analogs of receptor ligands are widely used in medicine. Compounds called agonists mimic the function of the natural ligand by binding to the receptor and inducing the normal response. Antagonists bind to the receptor but induce no response. Instead, they typically block binding and signaling by the natural ligand. Examples of an epinephrine agonist (isoproterenol) and antagonist (alprenolol) are shown in Fig. 15.11. Isoproterenol binds to bronchial smooth muscle cell epinephrine receptors with 10-fold higher affinity than epinephrine, and is used to treat asthma, etc. Alprenolol is a beta-blocker that binds to cardiac muscle cell epinephrine receptors, blocking epinephrine action and slowing heart contractions. It therefore helps treat cardiac arrhythmias and angina.
Structure of GPCRs G protein-coupled receptors (GPCRs) are the most numerous class of receptors in most eukaryotes. Receptor activation by ligand binding activates an associated trimeric G protein, which in turn interacts with downstream signal transduction proteins. All GPCRs are integral membrane proteins that have a common 7 transmembrane segment structure (Fig. 15.15). The hormone/ligand binding domain is formed by amino acids located on the external side of the membrane and/or membrane interior (Fig. 15.16a). Likewise in rhodopsin, its light absorbing chromophore 11-cis-retinal is located within the transmembrane segment interior of the protein. GPCRs interact with G proteins via amino acids in the C3 and C4 cytoplasmic regions.
G Protein Activation of Effectors The trimeric G protein cycle of activity in hormone-stimulated GPCR regulation of effector proteins is summarized in Fig. 15.17 (next slide). Initially, the G protein complex is tethered to the inner leaflet of the cytoplasmic membrane via lipid anchors attached to the Ga and Gg subunits. The trimeric GDP-bound form of the G protein is inactive in signaling. The binding of a hormone to the GPCR triggers a conformational change in the receptor (Step 1) which promotes its binding to the trimeric G protein (Step 2). Binding to the activated GPCR triggers the dissociation of GDP (Step 3). Subsequent binding of GTP to the Ga subunit activates it, and causes its dissociation from the receptor and the Gßg complex (Step 4). Ga-GTP then binds to the effector protein regulating its activity. The hormone eventually dissociates from the receptor (Step 5). Over time (often less than 1 min), GTP is hydrolyzed to GDP and Ga becomes inactive. It then dissociates from the effector and recombines with Gßg (Step 6). A hormone-bound GPCR activates multiple G proteins, until the hormone dissociates. Proteins known as regulators of G protein signaling (RGS) accelerate GTP hydrolysis by Ga decreasing the time-period during which Ga is active (not shown).
Trimeric G Proteins & Their Effectors There are 21 different Ga proteins encoded in the human genome. The G proteins containing these subunits are activated by different GPCRs and regulate a variety of different effector proteins (Table 15.1). The most common effectors synthesize second messengers such as cAMP, IP3, DAG, and cGMP. In the case of cAMP, a stimulatory Gs subunit activates adenylyl cyclase and cAMP production, whereas an inhibitory Gi subunit inhibits adenylyl cyclase and cAMP production.
GPCRs That Bind Epinephrine Epinephrine is a hormone that signals the "fight-or-flight" response. It elevates heart rate, dilates the airway, and mobilizes carbohydrate and lipid stores of energy in liver and adipose tissue. In the heart, liver, and adipose tissue, these effects are mediated via binding to ß1- & ß2-adrenergic GPCRs. Both ß-adrenergic GPCRs signal via Gas, which activates adenylyl cyclase and raises intracellular [cAMP]. The a2-adrenergic GPCR signals via Gai, decreasing adenylyl cyclase activity and intracellular [cAMP]. The a1-adrenergic GPCR is coupled to Gaq, which activates phospholipase C (PLC) and signaling via the IP3/DAG pathway (see below). a1-adrenergic GPCRs are present in the liver and blood vessels in peripheral organs. Binding to a1-adrenergic GPCRs stimulates glycogen breakdown in the liver, while blood flow to peripheral organs is decreased. Cholera toxin produced by Vibrio cholera, locks Gas-GTP in the active state, increasing [cAMP] in the large intestine, causing electrolyte and water loss. Pertussis toxin produced by Bordetella pertussis, locks Gai-GDP in the inactive state, increasing [cAMP] in the airway epithelium, causing mucus secretion into bronchial tubes, etc.
GPCRs that Regulate Ion Channels: Muscarinic Acetylcholine Receptor The neurotransmitter, acetylcholine (ACH) binds to two types of receptors known as the nicotinic and muscarinicacetylcholine receptors. The nicotinic receptor is itself a ligand-gated ion channel that opens on ACH binding. This receptor is located in the neuromuscular junctions of striated muscle. The muscarinic ACH receptor, is a GPCR found in cardiac muscle cells that is coupled to an inhibitory G protein (Fig. 15.20). The binding of ACH to this receptor triggers dissociation of Gai-GTP from Gßg, which in this case, directly binds to and opens a K+ channel. The movement of K+ down its concentration gradient to the outside of the cell, increases the positive charge outside the membrane, hyperpolarizing the cell. This results in the slowing of heart rate.
GPCRs that Regulate Ion Channels: Rhodopsin Rhodopsin is a light-activated GPCR found in the rod cells of the eye. Rhodopsin molecules are located within membrane disks in the outer segments of rod cells (Fig. 15.21). About 107 copies of rhodopsin occur per cell. Rod cells are important in capture of low intensity light having a broad range of wavelengths. Closely related color pigment receptors that respond to more limited regions of the visual spectrum (i.e., blue, green, & red light) are present in cone cells.
Mechanism of Rhodopsin Activation by Light Rhodopsin consists of the protein opsin bound to the visual pigment, 11-cis-retinal. Like other GPCR family members, rhodopsin is a 7-transmembrane segment protein. Rhodopsin signaling is initiated when the retinal chromophore absorbs a photon of light. Light absorption causes an electronic rearrangement and isomerization from 11-cis- to all-trans-retinal (Fig. 15.22). The isomerization triggers a conformational change in opsin, leading to activation of a bound G protein known as transducin (Gt). All-trans-retinal is released and recycled to 11-cis-retinal which later recombines with opsin.
Mechanism of Rhodopsin Signaling I The rhodopsin signal transduction pathway is shown in Fig. 15.23. Light absorption by rhodopsin triggers GTP/GDP exchange on the transducin Gat subunit, and dissociation of this trimeric G protein (Steps 1 & 2). Gat-GTP binds to and activates a cGMP phosphodiesterase, reducing intracellular cGMP level (Steps 3 & 4). This indirectly results in the closing of non-selective Na+/Ca2+ ion channels in the cytoplasmic membrane and hyperpolarization of the membrane potential (Step 6). This results in decreased release of neurotransmitter from the cells. Thus, light is perceived by the brain due to a decrease in nerve impulses coming from rod cells. Studies have shown that only 5 photons must be absorbed per human rod cell to transmit a signal. A single activated molecule of rhodopsin activates ~500 transducin molecules in a classic example of signal amplification.
Mechanism of Rhodopsin Signaling II Rhodopsin signaling must be rapidly shut down in order for the eye to detect rapid movement and other changes in objects in our surroundings. The shut down of signaling is accomplished in about 50 milliseconds, and involves several contributing processes. First, Gt-bound GTP is rapidly hydrolyzed.The hydrolysis of GTP by Gt is stimulated by a dimeric GAP protein consisting of the RGS9/Gß5 subunits (Step 7, preceding slide). Second, Ca2+-sensing proteins that detect a fall in intracellular Ca2+ stimulate the activity of guanylate cyclase, leading eventually to re-opening of ion channels (Fig. 15.23). Finally, the ability of activated rhodopsin to stimulate transducin is down-regulated by the phosphorylation of rhodopsin by rhodopsin kinase (Fig. 15.24). Signaling by triphosphorylated rhodopsin is completely blocked by the binding of a protein called arrestin.
Visual Adaptation Rod cell signaling actually is reduced after prolonged exposure to high light intensity. This is apparent as a time delay during which vision is compromised when we move from bright light to a dark room. The change in sensitivity of our eyes to high and low light levels is known as visual adaptation. The biochemical mechanism by which adaptation primarily occurs is shown in Fig. 15.25. In the dark, transducin molecule are transported to the outer rod segments, whereas arrestin molecules are transported elsewhere in the cell. In bright light, the distributions of transducin and arrestin are reversed. Through the distribution of these proteins, visual signaling is desensitized at high light levels and sensitized at low light intensities. Visual adaptation allows rod cells to perceive contrast over a 100,000-fold range of ambient light levels.
Synthesis and Hydrolysis of cAMP In the next few slides, we will cover signaling by the second messenger, cAMP. As shown in Fig. 15.26, cAMP is synthesized from ATP by the enzyme adenylyl cyclase. cAMP is broken down to AMP via the enzyme cAMP phosphodiesterase.
GPCRs that Regulate Adenylyl Cyclase Adenylyl cyclase is an effector enzyme that synthesizes cAMP. Ga-GTP subunits bind to the catalytic domains of the cyclase, regulating their activity. Gas-GTPactivates the catalytic domains, whereas Gai-GTPinhibits them. A given cell type can express multiple types of GPCRs that all couple to adenylyl cyclase. The net activity of adenylyl cyclase thus depends on the combined level of G protein signaling via the multiple GPCRs. In liver, GPCRs for epinephrine and glucagon both activate the cyclase. In adipose tissue (Fig. 15.27), epinephrine, glucagon, and ACTH activate the cyclase via Gas-GTP, while PGE1 and adenosine inactivate the cyclase via Gai-GTP.
Adenylyl Cyclase & Protein Kinase A Adenylyl cyclase is an integral membrane protein that contains 12 transmembrane segments (Fig. 15.28a). It also has 2 cytoplasmic domains that together form the catalytic site for synthesis of cAMP from ATP. One of the primary targets of cAMP is a regulatory kinase called protein kinase A (PKA), or cAMP-dependent protein kinase. PKA exists in two different states inside cells (Fig. 15.29a). In the absence of cAMP, the enzyme forms a inactive tetrameric complex in which 2 PKA catalytic subunits are non-covalently associated with 2 regulatory subunits. When cAMP concentration rises, cAMP binds to the regulatory subunits which undergo a conformational change, releasing the active catalytic subunits.
Regulation of Glycogen Degradation Glycogen is a polysaccharide that serves as the main store of glucose in many organisms. The liver stores glycogen for 1) release to the CNS during overnight fasting, and 2) release to skeletal muscle in response to epinephrine. Skeletal muscle stores glycogen for energy metabolism, which is accelerated by epinephrine. The reactions catalyzed by the key enzymes of glycogen synthesis (glycogen synthase) and degradation (glycogen phosphorylase) are shown in Fig. 15.31a.Epinephrine activates glycogen breakdown and blocks synthesis via activation of glycogen phosphorylase and inhibition of glycogen synthase. Epinephrine exerts these effects via raising cAMP levels through Gas-GTP signaling. The key target of cAMP is PKA. The activation of PKA leads to phosphorylation and activation of glycogen phosphorylase kinase and ultimately glycogen phosphorylase (left). In contrast, PKA inactivates glycogen synthase by phosphorylation. PKA also phosphorylates an inhibitor of phosphoprotein phosphatase, ensuring that protein phosphatase remains off (right). Hydrolysis of phosphates by protein phosphatase reverses the effects of PKA.
Activation of Gene Transcription by GPCR Signaling GPCRs regulate gene transcription by cAMP and PKA signaling. As shown in Fig. 15.32, cAMP-released PKA catalytic domains enter the nucleus and phosphorylate the CREB (CRE-binding) protein, which binds to CRE (cAMP-response element) sequences upstream of cAMP-regulated genes. Only phosphorylated p-CREB has DNA binding activity. p-CREB interacts with other TFs to help assemble the RNA Pol II transcription machinery at these promoters. In liver, glucagon signaling via this pathway activates transcription of genes needed for gluconeogenesis.
Down-regulation of GPCR/cAMP/PKA Signaling A number of events contribute to the termination of signaling by a GPCR. These include dissociation of the hormone from the receptor, hydrolysis of GTP by Ga, hydrolysis of cAMP via cAMP phosphodiesterase, and phosphorylation and “desensitization” of receptors by kinases such as PKA and ß-adrenergic receptor kinase (BARK). In addition, GPCRs can be removed from the membrane by vesicular uptake.
GPCRs That Activate Phospholipase C Another common GPCR signaling pathway involves the activation of phospholipase C (PLC). This enzyme cleaves the membrane lipid, phosphatidylinositol 4,5-bisphosphate (PIP2) to the second messengers, inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG) (Fig. 15.35). In this case, the Go and Gq G proteins conduct the signal from the GPCR to PLC. This is the pathway used in a1-adrenergic GPCR signaling in the liver. *
IP3/DAG Signaling Elevates Cytosolic Ca2+ The steps downstream of PLC that make up the IP3/DAG signalingpathway are illustrated in Fig. 15.36a. IP3 diffuses from the cytoplasmic membrane to the ER where it binds to and triggers the opening of IP3-gatedCa2+channels (Steps 3 & 4). Another kinase, protein kinase C (PKC) binds to DAG in the cytoplasmic membrane and is activated (Step 6). In liver, the rise in cytoplasmic [Ca2+] activates enzymes such as glycogen phosphorylase kinase, which phosphorylates and activates glycogen phosphorylase. Glycogen phosphorylase kinase is activated by Ca2+-calmodulin. In addition, PKC phosphorylates and inactivates glycogen synthase.
Nitric Oxide (NO)/cGMP Signaling A related signaling pathway involving phospholipase C operates in vascular endothelial cells and causes adjacent smooth muscle cells to relax in response to circulating acetylcholine (Fig. 15.37). In the NO/cGMP signaling pathway, the downstream target of Ca2+/calmodulin is nitric oxide synthase, which synthesizes the gas NO from arginine. NO diffuses into smooth muscle cells and causes relaxation by activating guanylyl cyclase and increasing [cGMP]. As a result arteries in tissues such as the heart dilate, increasing blood supply to the tissue. NO also is produced from the drug nitroglycerin which is given to heart attack patients and patients being treated for angina.