Download

1 / 41
510 likes | 1.4k Views
Osmoregulation. Ionic and osmotic balance. 2 /21. in multicellular organisms the interstitial fluid is the internal environment – composition resembles that of the ancient sea: high Na + , low K + , low Ca ++ and Mg ++

E N D
Ionic and osmotic balance 2/21 • in multicellular organisms the interstitial fluid is the internal environment – composition resembles that of the ancient sea: high Na+, low K+, low Ca++ and Mg++ • osmoregulatory organs maintain this environment isovolumic, isotonic, isoionic, isohydric, etc. • further task: removal of poisonous end products of metabolism (mostly NH3 from proteins) • marine invertebrates: osmotic pressure and ionic constitution in equilibrium with seawater • marine vertebrates: ion concentration is one third of seawater except in hagfish (Eptatretus - Cyclostomata), MgSO4, Cl- much lower; in sharks-rays ion concentration is lower, but osmotic pressure is maintained by urea • fresh water, terrestrial: ion concentration is one third; hyperosmotic to freshwater, hyposmotic to seawater Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Tab. 14-1.
Osmotic exchange 3/21 • plasma membrane separates fluids with different ionic composition, but equal osmotic pressure • epithelium separates fluids that are different in both respects • animals cannot isolate themselves from the environment: exchange of gases, absorption of nutritients; exception: Artemia salina • obligate and regulated osmotic exchange • obligate exchange depends on physical factors, that animals cannot readily regulate • regulated exchange compensates for changes caused by the obligate • only some parts of the epithelium participate in osmoregulation: gill, kidney, salt gland, enteric system
Obligate osmotic exchange 4/21 • occurs through the skin, respiratory epithelium and other epithelia in contact with environment • influencing factors: • gradient: determines direction of exchange; a frog sitting in a pond takes up water through the skin; a marine fish is loosing water in the sea, but gains NaCl; a freshwater fish is taking up water, but loosing salt through the gill • surface: small animal – relatively larger surface, faster exchange, e.g. dehydration • permeability: • transcellular and paracellular exchange (but: tight junction); • skin of amphibians, gill of fish have high permeability • skin of reptiles, desert amphibians, birds, mammals are much less permeable (leather containers), but mammals loose water through sweating • eating, metabolism, excretes: metabolic water is very important for desert animals, but also for marine mammals (seals put on weight eating fish, but loose weight burning fat when is feeding on invertebrates) • respiration: function of nose – condense water during exhalation – dropping nose in winter
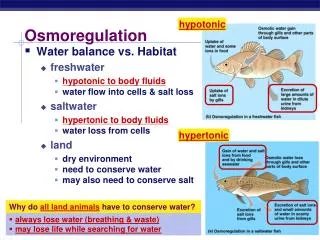
Osmotic regulation I. 5/21 • most vertebrates are strict osmoregulators - exception: shark, ray and hagfish • marine invertebrates are in equilibrium, other invertebrates, similar to vertebrates, are hyperosmotic in freshwater, hyposmotic in seawater • some of the invertebrates is conformer, others are osmoregulators • freshwater animals breathing from water • they are hyperosmotic: 200-300 mOsm; freshwater in general below 50 mOsm – water inflow, salt outflow • to compensate, they produce dilute urine, take up salt with food and through active transport (fish, frog), decrease the permeability of their skin • they do not drink freshwater Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-8.
Osmotic regulation II. 6/21 • marine animals breathing from water • invertebrates are in equilibrium • hagfish: Ca++, Mg++, SO42- regulation only • shark-ray osmotic equilibrium due to urea accumulation, excess salt removed by rectal glands • fish are loosing water, drinking seawater; excess salt removed through the gill (chloride cells) • marine animals breathing air • loosing water through respiratory epithelium and other epithelia • marine reptiles and birds are drinking seawater – cannot produce strongly hyperosmotic urine (just as fish) – excess salt removed through salt glands • marine mammals do not drink seawater, water is taken in with food and produced metabolically, urine is hyperosmotic • lion seal males spend 3 months on the beach without eating or drinking; seal pups do similarly for 8-10 weeks while mother is on the sea Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-8.
Osmotic regulation III. 7/21 • terrestrial animals breathing air • if freshwater is available then water lost through breathing can be replenished by drinking, salt loss (urine, faces, sweating) compensated from food – sparing is important • problem of shipwrecked sailors: kidney can remove 6 g Na+/l urine, seawater contains 12 g Na +/l – drinking of seawater leads to salt gain • desert animals have two problems: heat and lack of water • kangaroo rat: remains in cool burrow during daytime, active only during the night; gaining water metabolically, water condensation in nose • camel: cannot hide in burrow; when dehydrated do not sweat, body temperature raises changing between 35-41° C; hyperosmotic or no urine, urea stored in tissues; faces dry Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-8.
Overview 8/21
Water compartments 9/21 • human body contains 60% water on average, differences between male-female, young-old • distributed in different compartments • intracellularly 2/3, extracellularly 1/3 • of the extracellular water: 3/4 interstitially, 1/4 in blood plasma • barriers and, transport rules are already known • measurement of volumes applying dilution principle: Evans-blue, inulin, tritiated water • homeostasis is very important . cholera, diarrhea - dehydration, working by a furnace in tropical areas – water poisoning, severe burns – dehydration due to loss of skin • the most important regulator in humans is the kidney, behavioral regulation is also important – metabolic water is limited
Human kidney 10/21 • osmoregulatory organs always contain transport epithelium (skin, gill, kidney, gut) : polarized - apical (luminal, mucosal) and basal (serosal) surfaces are different • capacity of the transport epithelium is increased by its special structure: tubular organization • functioning of the mammalian kidney is well known – though it does not represent all types of vertebrate kidneys • 0,5% of body weight, 20-25% of cardiac output • cortex, medulla, renal pyramid, renal pelvis, ureter, urinary bladder, urethra • volume of urine is 1 l daily, slightly acidic (pH 6), composition, volume changes with the food and the requirements of the water homeostasis - beer, Amidazophen, etc. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-13.
The nephron 11/21 • functional unit of human kidney is the nephron • afferent and efferent arterioles, in between glomerulus; Bowman capsule, proximal tubule, loop of Henle, distal tubule, collecting duct • most of the nephrons (85%) are cortical, the rest juxtamedullary (15%) nephron • steps in the formation of urine: • ultrafiltration • reabsorption • secretion • the kidney is very important in pH regulation • the kidney removes ammonia formed during the decomposition of proteins Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-14.
Ultrafiltration 12/21 • in the kidney 15-25% of water and solutes is filtrated, 180 l daily – proteins and blood cells remain • filtration depends on: • the hydrostatic pressure between the capillaries and the lumen of the Bowman capsule: 55-15 = 40 mmHg • the colloid osmotic pressure of the blood: 30 mmHg – effective filtration pressure 40-30 = 10 mmHg • the permeability of the filter: fenestrated capillaries, basal membrane (collagen + negative glycoproteins), podocytes (filtration slits between pedicels) • voluminous blood supply due to the relatively low resistance – afferent arteriole is thick and short – high pressure in the glomerulus • regulation of the blood flow: basal miogenic tone, paracrine effect of juxtaglomerular apparatus, sympathetic effect (afferent arteriole, glomerulus, podocyte) Berne and Levy, Mosby Year Book Inc, 1993, Fig. 41-7 Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-20. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-19. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-18
Clearance 13/21 • clearance of a substance is the volume of plasma that is completely cleaned from the given substance in the kidney in every minute VUCP = VUthat isC = ------P C - clearance, P – concentration in plasma, V – volume of urine in 1 minute, U – concentration in urine • clearance of a substance that is neither reabsorbed nor secreted (e.g. inulin) equals the glomerulus filtration rate :GFR • clearance of a substance that is not only filtrated, but completely secreted as well (e.g. PAH) equals the renal plasma flow :RPF • knowing the hematocrit, renal blood flow (RBF) can be calculated
Tubular reabsorption I. 14/21 • 180 l primary filtrate is produced every day, but only 1 l is excreted, of 1800 g filtrated NaCl only 10 g remains in the urine • the process of reabsorption has been successfully examined since the 1920’s using the method of micropuncture • role played by the subsequent sections of the tubules: • proximal tubule • 70% of Na+ is reabsorbed by active transport, Cl- and water follow passively, obligate reabsorption • filtrate is isosmotic, but concentration of substances that are not reabsorbed increases 4-fold • on the apical membrane of epithelial cells microvilli • virtually all filtrated glucose and amino acids are reabsorbed using Na+ dependent symporter • tubular maximum for glucose: below 1.8 mg/ml complete reabsorption (normal value: 1.0 mg/ml), above 3.0 mg/ml linear increase – sugar in urine in diabetes • Ca++, phosphate and other electrolytes are reabsorbed as needed – see later Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-25.
Tubular reabsorption II. 15/21 • descending part of Henle’s loop • no microvilli, few mitochondria – no active transport • low permeability for NaCl and urea, high for water • thin ascending part of Henle’s loop • no microvilli, few mitochondria – no active transport • low permeability for water, high for NaCl • thick ascending part of Henle’s loop • active reabsorption of Na+ • low water permeability • distal tubule • active reabsorption of Na+, and passive reabsorption of water • K+, H+ and NH3 transport as needed – see later (pH regulation) • transport is regulated by hormones – facultative reabsorption • collecting duct • active reabsorption of Na+ at the cortical part, high urea permeability in the internal medullary part • regulated water permeability (ADH) Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-24.
Tubular secretion 16/21 • several substances are secreted from the plasma to the tubule in the nephron – best examined: different electrolytes (K+, H+, NH3) organic acids and bases • active transport – recognizes substances conjugated with glucuronic acid in the liver • K+ is reabsorbed in the proximal tubule and Henle’s loop (Na/2Cl/K transporter) • if K+ concentration is too high, secretion in the distal tubule depending on aldosterone and coupled to Na+ reabsorption - K+ acts directly on aldosterone, Na+ through renin-angiotensin • conflicting demands: insulin secretion is induced by high K+ - excess K+ is taken up by adipose tissue • secretion of H+ and NH3 serves pH regulation Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-28.
pH regulation I. 17/21 • normal pH 7.4 – 7.35 acidosis, 7.45 alkalosis • normal functioning is possible between 7.0-7.8 • regulation: buffer systems, respiration, kidney • Henderson-Hasselbalch equation: [A-] pH = pK + log ------ [HA] • for CO2 two equations; following rearrangement: • [HCO3-] pH = pK’ + log ------ [αCO2] • - solubility of CO2 • pK’ = 6.08, i.e. not good buffer at normal pH – as CO2 and HCO3- can be easily modified (respiration, kidney) • plasma proteins (14-15% Hgb, 6-8% other) pK is same as blood pH – good buffers • phosphate concentration low, negligible effect
pH regulation II. 18/21 • respiratoryalkalosis and acidosis: caused by hyper-, or hypoventilation • metabolic alkalosis – e.g. Cl- loss because of vomiting • metabolic acidosis – anaerobic energy production, ketosis in diabetes mellitus • in the first case: kidney compensates, in the second: breathing (short-term) and the kidney (long-term) • proximal tubule, Henle’s loop: Na+/H+ exchanger, distal tubule, collecting duct: HCO3- uptake through A-cells • in distal tubule and collecting duct: HCO3- secretion through B-cells • in acidosis HCO3- level is low in the filtrate: NH3 secretion – binds to H+, NH4+ cannot go back, H+ secretion increases Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-30. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-29.
Hyperosmotic urine 19/21 • birds and mammals can produce hyperosmotic urine - water reabsorption in the collecting duct due to osmotic pressure differences • common characteristic: Henle’s loop, the longer the loop, the more concentrated the urine – very long in kangaroo rat • pressure difference is achieved through the counter-current principle • Na+ transport in the ascending part of the Henle’s loop – do not enter the descending part, but attracts water leading to the same result • in addition, urea present in high concentration because of the reabsorption of water, can only leave the tubule in the internal medulla • osmotic pressure increases from the cortex to the medulla • blood supply to the tubules (vasa recta) is running in parallel to the Henle’s loop , does not decrease the osmotic gradient Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-32. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, SL. 14-2. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-32.
Regulation of the kidney 20/21 • granular cells in the juxtaglomerular apparatus produce renin in response to a decrease in blood pressure or NaCl delivery to the distal tubule • renin cuts off angiotensin I (10 amino acids) from angiotensinogen (glycoprotein) • converting enzyme (mostly in the lung) cuts off 2 amino acids from angiotensin I – angiotensin II • angiotensin II enhances aldosterone secretion in the adrenal gland, increases blood pressure through vasoconstriction and increases ADH production • aldosterone increase Na+ reabsorption through 3 different ways: facilitation of the pump, ATP production, increased apical Na+ permeability • ADH producing cells detect blood pressure and osmolality and are sensitive to alcohol • atrial natriuretic peptide (ANP) – released in the atria when venous pressure increases - inhibits renin, aldosterone, ADH production Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-35. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-26. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-27.
Nitrogen removal 21/21 • part of the digested amino acids are reused, amino groups from the others have to be removed as NH3 and NH4+ are poisonous • three forms: ammonia, urea, uric acid • ammonia • poisonous – huge volume is needed to provide low concentration in the cell and high outward gradient • 0.5 l water/1 g nitrogen • fish, aquatic invertebrates, mammals in low amount • transport in the form of glutamine from the liver to the kidney • urea • less poisonous, 0.05 l water/1 g nitrogen • synthesis requires ATP • vertebrates, except fish, synthesize urea in the ornithine-urea cycle, fish and invertebrates from uric acid • hominoids cannot metabolize uric acid (from nucleic acids) – can accumulate and lead to gout • uric acid • low solubility: 0.001 l water/1 g nitrogen • white precipitate - guano in birds (uric acid, guanine) • fish, reptiles, terrestrial arthropods Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-49, 50.
Extracellular ion concentrations Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Tab. 14-1.
Osmoregulation in animals Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-8.
Structure of mammalian kidney Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-13.
Structure of a nephron Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-14.
Glomerular filtration Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-18
Podocytes of the capsule Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-19.
Podocytes of the capsule - EM Berne and Levy, Mosby Year Book Inc, 1993, Fig. 41-7
Juxtaglomerular apparatus Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-20.
Na+ reabsorption Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-25.
Processes of reabsorption Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-24.
Mechanism of K+ secretion Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-28.
pH regulation Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-29.
Release of ammonia Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-30.
Counter-current principle Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, SL. 14-2.
Mechanism of urine concentration Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-32.
Osmotic pressure in the kidney Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-32.
Renin-angiotensin system Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-26.
Actions of aldosterone Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-27.
Regulation of ADH secretion Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-35.
Methods of nitrogen release Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 14-49, 50.