Download

1 / 22
220 likes | 399 Views
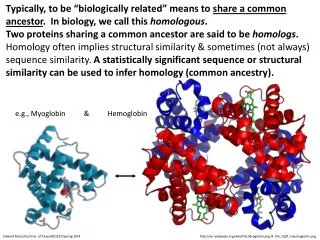
Myoglobin- Key Properties. An O 2 transport protein in muscle A globular soluble protein, 151 residues (16 kDa) 8 a -helices (A,B,C,…..H)- first protein crystal structure! Contains a heme prosthetic group Fig. 7-1.

E N D
Myoglobin- Key Properties • An O2 transport protein in muscle • A globular soluble protein, 151 residues (16 kDa) • 8 a-helices (A,B,C,…..H)- first protein crystal structure! • Contains a heme prosthetic group • Fig. 7-1
The Heme Prosthetic GroupThe O2 carrier in Myoglobin and Hemoglobin • Protoporphyrin with Fe(II) • Covalent attachment of Fe via His F8 side chain • Additional stabilization via hydrophobic interaction • Fe(II) state is active, Fe(III) [oxidized] • Fe(II) atom in heme binds O2 Figs. 7-2 & 7-3
Binding of O2 to Heme • Binding of O2 to a free heme group is irreversible • Enclosure in a protein allows reversible binding • O2 has only limited solubility (1 X 10-4 M) in water • Solubility problem overcome by binding to proteins • Also increases diffusion • • Binding of O2 alters heme electronic structure • Causes changes in heme electronic spectrum (Vis) • Bright scarlet color of blood in arteries • Dark purple color of blood in veins
Carbon Monoxide Poisoning • Heme Fe(II) binds many other small molecules with structures similar to O2 including: CO, NO, H2S • O2 is actually a fairly weak binder relative to these other molecules, particularly CO. [Essential for Biology] • • When exposed to CO, even at low concentrations, O2 transport proteins will be filled with CO limiting their vital O2 capacity.
Myoglobin O2 Binding Properties Mb + O2 MbO2 • Dissociation constant, Kd = [Mb] [O2] / [MbO2] Quick Review [from enzymes] • If Kd (dissociation) is higher, binding is weaker • If binding curve is less steep, binding is weaker • Half-saturation is the point where [O2] = Kd: Kd = [Mb] [O2] / [MbO2] = [Mb] Kd / [MbO2] Kd / Kd = [Mb] / [MbO2] = 1 …… [Mb] = [MbO2]
Myoglobin O2 Binding Properties Mb + O2 MbO2, Kd = [Mb] [O2] / [MbO2] • To think about O2 transport, express binding in terms of the fraction of occupied binding sites, i.e. Fractional Saturation (YO2): YO2 = [MbO2] / ([Mb] + [MbO2]) • Substituting with Kd, allows understanding of transport properties in terms of O2 availability: YO2 = [O2] / (Kd + [O2])
Myoglobin O2 Binding Properties • YO2 = [O2] / (Kd + [O2]) • • O2 is a gas: express [O2] as partial pressure (pO2) • [Common to call pO2oxygen tension] • YO2 = pO2 / (Kd + pO2) • As pO2 increases, binding increases • Binding curve is hyperbolic, not linear (just like enzymes!!!) [Draw] • Low pO2 little binding to protein • High pO2 saturation of protein • At half-saturation pO2 = Kd, field uses term P50 • P50(myoglobin) = 2.8 torr / pO2 = 10-100 torr in-vivo!!
Hemoglobin- Key Properties • Ubiquitous O2 transport protein • A globular soluble protein, 2X2 chains (164 kDa) • a and b chains 44% identical • All helical secondary structure (like myoglobin) • abab quaternary structure • a-subunit 141 residues • b-subunit 146 residues • Extensive contacts between subunits • Mix of hydrophobic, H-bond, and ionic interactions • a1b1 (a2b2)- 35 residues, a1b2 (a2b1)- 19 residues
Structure of Hemoglobin Heme a2 b1 b2 a1 Inter-subunit contacts
Hemoglobin O2 Binding Properties Hb + nO2 Hb(O2)1 + Hb(O2)2 + Hb(O2)3 + Hb(O2)4 Kd = [Hb] [O2]n / [Hb(O2)1 + Hb(O2)2 + Hb(O2)3 + Hb(O2)4] [How’s that for complicated!?!?!] • To think about O2 transport, express binding in terms of the fraction of occupied binding sites, i.e. Fractional Saturation (YO2): YO2 = [Hb(O2)1 + ….] / ([Hb] + [Hb(O2)1 + ….])
Hemoglobin O2 Binding Curve • Binding curve is sigmoidal • Artery: high pO2, loading of protein • Vein: lower pO2, unloading from protein • P50(hemoglobin) = 26 torr, adjusts as needed!! *Drastic change in pO2 over physiological range*
Cooperativity in Binding O2 The sigmoidal shape is a consequence of the 4 subunits of hemoglobin "cooperating" in the binding of O2. • As pO2 increases and [O2] increases, increasing probability that at least 1 subunit has bound O2. Binding of O2 to a subunit INCREASES the probability that empty subunits will be able to bind an O2!! • As pO2 increases even further, the probability that remaining binding sites will have O2 bound increases. • Eventually, a plateau is reached: when most hemoglobins are filled there are few sites left to bind to, so not much increase, even if the pO2 is very high.
The Hill Equation • Simplification is required to solve binding equation: assume O2 binding cooperativity is infinite • Binding curve can be expressed in terms of [O2]: YO2 = [O2]n / (Kd + [O2]n) • O2 is a gas: substitute pO2 for [O2] and use Kd = P50: YO2 = (pO2)n / (P50 + (pO2))n This expression for the degree of saturation of Hb is known as the Hill Equation • The quantity n is called the Hill Constant
Interpretation of the Hill Equation • YO2 = (pO2)n / (P50 + (pO2))n • n: the degree of cooperativity in ligand binding • n = 1: non-cooperative • n > 1: positive cooperativity implies binding of a ligand increases affinity for next ligand. • n < 1: negative cooperativity implies binding of a ligand decreases affinity for next ligand. • Rearranging the Hill equation allows Hill Plots to be made to facilitate analysis of binding curves.
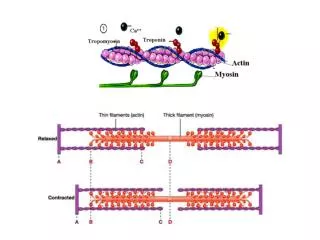
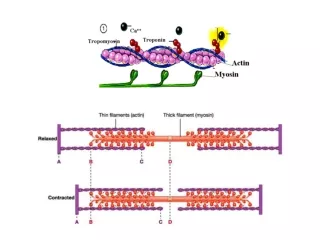
Binding of O2 to the Heme Changes the Whole Structure of Hemoglobin R state T state b chains further apart Shifts at the ab interfaces
The T to R State Transition • Binding of O2 causes a series of shifts in all subunits • Change in heme structure upon binding O2 • Since His F8 is covalently attached, all of F helix shifts • Reorganization of helix alters tertiary structure, which in turn alters the quaternary structure- 4 chains behave as a single cooperative structural unit • Changes in packing of hydrophobic side chain • Changes in pairing of charged side chains • The change in conformation of Hemoglobin from the T to the R state increases O2 affinity at ALL sites
Allosteric Effectors • The R or T state can be stabilized by the binding of ligands other than O2. • H+. Lower pH favors the T state which causes Hb to release bound O2. This is known as the Bohr Effect. • CO2. Release of CO2 lowers pH via conversion to HCO3-: CO2 + H2O HCO3- + H+. Reinforces Bohr Effect • Bisphosphoglycerate (BPG). Regulation of activity via binding more strongly to T state, helps to release O2. Increase in levels of BPG helps adaptation to high altitude- faster than making more hemoglobin. Also important in hypoxia diseases (e.g. anemia)
Towards a More Complete Picture Model for disucssion HEMOGLOBIN at the pH (~7.6) found in the lungs. HEMOGLOBIN at the pH (~7.2) found in peripheral tissues. MYOGLOBIN in muscle (a peripheral tissue).
Path of O2 Flow 1. O2 diffuses from the alveoli of the lungs into the capillaries of the bloodstream then into the red blood cells 2. In the red blood cells, O2 binds to hemoglobin. 3. In parallel, CO2 diffuses from blood into the alveoli. 4. The lower concentration of dissolved CO2 in the blood causes lower pH (~7.6) in lungs than in the peripheral tissues (~pH 7.2) where CO2 is being actively released. A. High pO2 / high pH
Why O2 Transport Works • 5. Red blood cells (containing O2-hemoglobin) carried to the peripheral tissues. B. pO2 decreases because O2 USED by the tissues. C. Blood plasma becomes more acidic (lower pH) because CO2 is released. • The combination of lower pO2 and pH in the peripheral tissues causes a large decrease in O2 saturation. • O2 is released by hemoglobin!!!! Note: changes in pO2 and pH are small!
Why Myoglobin in Muscle? • Under resting conditions, O2 saturation is at point X on the green curve • Small changes in pO2 and pH have very little effect on saturation • During extremely vigorous exercise, heart pumps blood fast and breathing is rapid to increase the intake of O2 . Also, pH is lowered. • Eventually, transport not fast enough to meet needs, i.e. pO2 lowered because O2 is used faster than it can be replenished. [Hemoglobin now no help!] • Under extreme conditions, shift from point X to Y: saturation of the myoglobin is lowered = release of O2.
Defects from Hemoglobin Mutations • Weakened heme binding. • Disruption of secondary structure. • Disruption of quaternary structure. • Defective oxygen transfer. • Altered affinity for oxygen. • Oxidation of Fe(II) to Fe(III). • Aggregation in the T state (Hemoglobin S). Sickle cell anemia results from aggregation of Hb into insoluble fibers causing mishapen blood cells that cannot pass through capillaries and block blood flow to tissues.