Download

1 / 43
440 likes | 616 Views
Viruses and Cancer. Orla Sheils Department of Histopathology. Carcinogenesis. What Causes Cancer?. Carcinogens Age genetic make up immune system diet day-to-day environment Viruses. Age. Diet. Factors in Carcinogenesis. Chromosomes/DNA.

E N D
Viruses and Cancer Orla Sheils Department of Histopathology
What Causes Cancer? • Carcinogens • Age • genetic make up • immune system • diet • day-to-day environment • Viruses
Factors in Carcinogenesis Chromosomes/DNA
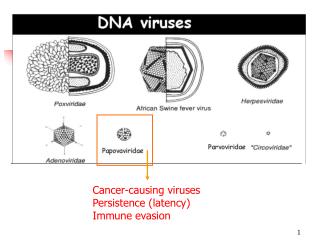
In addition to chemicals and radiation, a few viruses also can trigger the development of cancer. In general, viruses are small infectious agents that cannot reproduce on their own, but instead enter into living cells and cause the infected cell to produce more copies of the virus. Like cells, viruses store their genetic instructions in large molecules called nucleic acids. In the case of cancer viruses, some of the viral genetic information carried in these nucleic acids is inserted into the chromosomes of the infected cell, and this causes the cell to become malignant.
Viral Oncology –historical perspective • First tumour viruses described at start of 20th Century • Retroviruses • Avian leukaemia virus - Ellerman & Bang • Avian sarcoma virus –Rous • Ludwig Goss 1950s - retroviruses cause tumours in mice • William Jarrett – feline leukaemia virus
Viruses Viruses contribute to development of some cancers. Typically, the virus can cause genetic changes in cells that make them more likely to become transformed. These cancers and viruses are linked • Cervical cancer and the genital wart virus, HPV • Primary liver cancer and the Hepatitis B virus • T cell leukaemia in adults and the Human T cell leukaemia virus
HTLV-1 • naturally infects CD4+ T lymphocytes and can be transmitted between close contacts through blood transfer or from mother to infant through cells in breast milk. • In most cases the infection is harmless. However, as many as 1 in 20 infected individuals eventually develop a type of adult T cell leukaemia in which every tumour cell carries a clonally integrated HTLV1 provirus. • HTLV1 differs from the standard 'chronically oncogenic' and 'acutely oncogenic' retroviruses in its mechanism of action; • it appears to drive cell growth through expression of a particular viral protein, Tax, in latently-infected cells
Mode of action • Tax can transactivate expression of a number of key cellular genes that enhance cell growth. • The best examples are the genes encoding • interleukin 2 (a T cell growth factor) and • the interleukin 2 receptor (a molecule that allows cells to respond to the growth factor). • As a consequence, the infected cells not only make their own growth signals, but also respond to them
Mode of action • HTLV1 induces a weak growth transformation of T cells in the laboratory but, in the body, is probably never sufficiently strong to induce T cell leukaemia on its own. • BUT, a virally infected cell in which growth controls have even partly broken down, is more susceptible to further genetic accidents. • During persistent infection a gradual build-up of HTLV1-positive T cells which have accumulated additional genetic changes may occur. • Eventually this can lead to selection and outgrowth of a fully malignant, HTLV1-positive clone • At this stage malignant cell growth can occur in the absence of tax gene expression.
Epstein Barr Virus • Epstein-Barr virus (EBV), also called Human herpesvirus 4 (HHV-4), is a virus of the herpes family (which includes Herpes simplex virus and Cytomegalovirus), • one of the most common viruses in humans. • Most people become infected with EBV, • often asymptomatic • but commonly causes infectious mononucleosis. • It is named after Michael Epstein and Yvonne Barr, who together with Bert Achong discovered the virus in 1964.
On infecting the B-lymphocyte, the linear virus genome circularises and the virus subsequently persists within the cell as an episome. • The virus can execute several distinct programmes of virally-encoded gene expression • broadly categorised as being lytic cycle or latent cycle. • The lytic cycle or productive infection results in staged expression of a host of viral proteins with the ultimate objective of producing infectious virions. Formally, this phase of infection does not inevitably lead to lysis of the host cell as EBV virions are produced by budding from the infected cell. • The latent cycle programmes are those that do not result in production of virions.
EBV-associated malignancies • The strongest evidence linking EBV and cancer formation is found in Burkitt's lymphoma and Nasopharyngeal carcinoma
Burkitts Lymphoma • a type of Non-Hodgkin's lymphoma • most common in equatorial Africa • co-existent with the presence of malaria. • Malaria infection causes reduced immune surveillance of EBV immortalised B cells, so allowing their proliferation. This proliferation increases the chance of a mutation to occur. Repeated mutations can lead to the B cells escaping the body's cell-cycle control, allowing the cells to proliferate unchecked, resulting in the formation of Burkitt's lymphoma. Burkitt's lymphoma commonly affects the jaw bone, forming a huge tumour mass. It responds quickly to chemotherapy treatment, namely cyclophosphamide, but recurrence is common. • Other B cell lymphomas arise in immunocompromised patients such as those with AIDS or who have undergone organ transplantation with associated immunosuppression. • Smooth muscle tumours are also associated with the virus.
Nasopharyngeal carcinoma • found in the upper respiratory tract, most commonly in the nasopharynx, and is linked to the EBV virus. • It is found predominantly in Southern China and Africa, due to both genetic and environmental factors. It is much more common in people of Chinese ancestry (genetic), but is also linked to the Chinese diet of a high amount of smoked fish, which contain nitrosamines, well known carcinogens (environmental).
When EBV infects B-lymphocytes in vitro, lymphoblastoid cell lines eventually emerge that are capable of indefinite growth. • The growth transformation of these cell lines is the consequence of viral protein expression. • EBNA-2, EBNA-3C and LMP-1 are essential for transformation while EBNA-LP and the EBERs are not. • The EBNA-1 protein is essential for maintenance of the virus genome
EBNA-2 is the main viral transactivator, switching transcription from the Wp promoters used during initial infection to the Cp promoter. • Together with EBNA-3C, it also activates the LMP-1 promoter. It is known to bind the host RBP-Jκ protein that is a key player in the Notch pathway. EBNA-2 is essential for EBV-mediated growth transformation. • EBNA-3A/EBNA-3B/EBNA-3C also bind the host RBP-Jκ protein. • EBNA-3C is also a ubiquitin-ligase and has been shown to target cell cycle regulators like pRb.
Kaposi's sarcoma • form of skin cancer that can involve internal organs. It most often is found in patients with acquired immunodeficiency syndrome (AIDS), and can be fatal.
K.S. • Kaposi's sarcoma (KS) was once a very rare form of cancer, primarily affecting elderly men of Mediterranean and eastern European background (tumours on lower legs), until the 1980s, when it began to appear among AIDS patients. • AIDS-related KS, emerged as one of the first illnesses observed among those with AIDS. Unlike classic KS, AIDS-related KS tumours generally appear on the upper body, including the head, neck, and back. The tumours also can appear on the soft palate and gum areas of the mouth, and in more advanced cases, they can be found in the stomach and intestines, the lymph nodes, and the lungs.
Kaposi Sarcoma and HHV8 • Studies in 2000 showed that HHV-8 was the culprit behind KS. • It does not work alone. • In combination with a patient's altered response to cytokines (regulatory proteins produced by the immune system) and the HIV-1 transactivating protein Tat which promotes the growth of endothelial cells, HHV-8 can then encode interleukin 6 viral proteins, specific cytokines that stimulate cell growth in the skin. • This becomes KS.
HHV-8 • HHV-8 destroys the immune system further by directing a cell to remove the major histocompatibility complex (MHC-1) proteins that protect it from invasion. • These proteins are then transferred to the interior of the cell and are destroyed. • This leaves the cell unguarded and vulnerable to invaders which would normally be targeted for attack by the immune system.
Global burden of cervical cancer Age standardised incidence rates per 100,000
Genomic Map of HPV (von Knebel Doberitz/European Journal of Cancer 2002; 38: 2229-2242).
p16(INK4a) p16(INK4a) MCM5 Geminin Pathways of Cell Cycle Regulation in Cervical Cancer p16(INK4a)
The Hepatitis B Virus (HBV) • HBV is a mostly double-stranded DNA virus in the Hepadnaviridae family. • HBV causes hepatitis in human. • The HBV genome has four genes: pol, env, pre-core and X that respectively encode the viral DNA-polymerase, envelope protein, pre-core protein (which is processed to viral capsid) and protein X. • The function of protein X is not clear but it may be involved in the activation of host cell genes and the development of cancer.
Consequences of HBV Infection • HBV causes acute and chronic hepatitis. • The chances of becoming chronically infected depends upon age. • About 90% of infected neonates and 50% of infected young children will become chronically infected. • In contrast, only about 5% to 10% of immunocompetent adults infected with HBV develop chronic hepatitis B.
Hepatocellular carcinoma • cancer that arises from hepatocytes, the major cell type of the liver. • Hepatocellular carcinoma is one of the major cancer killers. • It affects patients with chronic liver disease who have established cirrhosis, and currently is the most frequent cause of death in these patients. • The main risk factors for its development are hepatitis B and C virus infection, alcoholism and aflatoxin intake.
HBV DNA integration • Should be viewed as a dynamic process. • Longterm chronic inflammation, associated with increased liver cell proliferation, induces several rearrangements of the integrated viral sequences (Hessein et al., 2005). • Deletions of part of the viral genome as well as more complex rearrangements are frequently observed (Wang et al., 2004b). • Hepatitis B virus insertion can induce chromosomal deletions at the HBV integration sites. • Transpositions of the viral sequences, together with the flanking cellular sequences from one chromosome to another, have been recently confirmed
integrated HBV can generate chromosomal instability • it has been suggested that viral DNA sequences encompassing the encapsidation signal may exhibit intrinsic recombinogenic activity via binding to a putative ‘recombinogenic’ cellular protein
HBV integration The insertion of HBV genome in cellular genes frequently targets genes that regulate key cellular pathways. The figure illustrates that HBV targets a variety of genes controlling various steps of cellular signalling, cell proliferation and viability
HBx Targets modulated by HBx -
Targets of HBx Schematic representation of the different targets of HBx. The figure shows the cytoplasmic, mitochondrial and nuclear transduction cascades activated by HBx.