Download
1 / 2
20 likes | 147 Views
fMRI Repetition Effects of Masked Picture Priming Marianna Eddy 1 , David Schnyer 2 , Annette Schmid 1 , Phillip J. Holcomb 1 1 Tufts University, 2 Boston University School of Medicine. INTRODUCTION

E N D
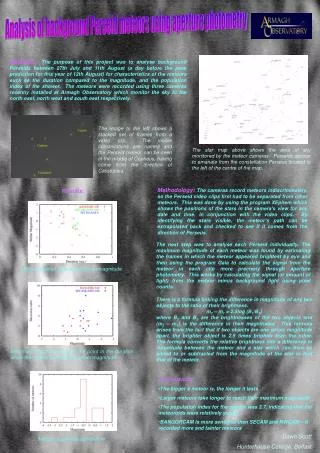
fMRI Repetition Effects of Masked Picture PrimingMarianna Eddy1, David Schnyer2, Annette Schmid1, Phillip J. Holcomb11Tufts University, 2Boston University School of Medicine • INTRODUCTION • A previous ERP study (Eddy et al., 2004) of masked picture priming revealed two important ERP effects of repetition – one in the range of the N400 and an earlier effect in the range from 100-300 msec. However, the spatial resolution of ERPs is poor, so the current study used an event-related fMRI design in order to better determine the neuroanatomical underpinnings of masked picture priming. • Previous studies of masked word priming have revealed effects of repetition suppression in the fusiform cortex (e.g., Dehaene et al., 2001; 2004). Unmasked picture priming studies have also found repetition suppression in the fusiform cortex (Buckner et al., 1998; Koutstaal et al., 2001). Finally both the word and picture studies have also revealed exemplar specific effects. For example, Koutstaal et al. (2001) found less repetition reduction in the right fusiform than the left fusiform for repeated different picture exemplars. • Problem: in standard priming techniques used in picture studies so far, the participants are • consciously aware of the prime and can use strategies that may interfere with observing the • phenomenon of interest. • Solution: masked-priming, “covering” the prime and making participants unaware of what they • have seen. • HYPOTHESES • Similar repetition effects will be seen with masked pictures as have been seen with masked words and unmasked pictures (i.e., repetition suppression in the fusiform cortex), • An effect of different exemplars (i.e., a semantic effect) may be observed that will differ between the left and right fusiform. RESULTS The masking paradigm worked well (participants reported that they knew there was something in the prime position, but where unable to identify it). They responded 97% of time to food targets, but only 15% to food primes Mean values from timepoints 4 and 6 were extracted from the anterior and posterior portions of the fusiform gyrus as well as the posterior inferior temporal gyrus for each individual subject. Mean percent signal change, baselined to the average of the 4 sec. prestimulus period, was calculated for the average of timepoints 4-6 and entered into a repeated measures ANOVA. Using words in a similar masked priming paradigm Dehaene et al. (2004) noted anterior – posterior differences in the fusiform gyrus. In the current study, the fusiform gyrus was divided into anterior – posterior portions by dividing each individual subject’s fusiform in half on the inflated spherical surface (See Figure 5). • DATA ANALYSIS • The fMRI data was analyzed using Freesurfer (FS-FAST stream). Data was motion corrected, smoothed with a 6 mm Gaussian filter, and intensity normalized before estimation of the event-related responses was accomplished using a fixed impulse response model. • High resolution MPRAGE images were segmented and cortical regions defined using a automated surface-based analysis that transformed each individual’s brain into a common spherical coordinate system (see Dale et al., 1999; Fischl et al., 1999 for details of procedure). Each individual’s fMRI results were aligned with their structural images and transformed into spherical coordinates. • Whole brain analysis (see Figure 2 for brain coverage) • a random effects group average was computed for voxels whose hemodynamic response fit a predesignated gamma function and showed greater activation in the 3 priming conditions (primed, unprimed, exemplar) versus the control picture (see Figure 3, left). • Additionally, direct pairwise comparisons between priming conditions were computed from random effects group averages at timepoint 4 (Figure 3, right). • Regions of Interest (ROI) analysis • the comparison of the 3 priming conditions greater than the control, was used as a functional constraint for the anatomically defined ROIs (see Figure 5), thresholded at p=0.001. Figure 3: Random Effects Group Average Fusiform Post ITG Fusiform Figure 5: Anterior/Posterior Fusiform Gyrus and Posterior Inferior Temporal Gyrus Fusiform Post ITG Fusiform Post ITG Post ITG Single Representative Subject
METHODS • Masked Picture Priming – Semantic Categorization Task • 11 participants (5 females, mean age = 20 years) • Participants were presented pictures projected onto a screen and instructed to respond when they detected food items. • 60 trials of each condition (see Figure 1) • 60 trials of probe pairing (30 with food item in prime position • 30 with food item in target position) • Control condition (60 trials) and null event of fixation (20% of trials) • 8 counterbalanced lists • Primes were forward and backward masked with a pixelated image • fMRI data was collected on a 3T Siemens Trio system using an 8-channel head coil • The slice prescription was chosen to maximize the signal in the temporal lobes (see Figure 2) • TR = 2000 ms, TE = 30 ms, 23 axial slices, 2 x 2 x 2.5 mm, 0.2 mm interslice gap (96x96 acquisition matrix) • Fusiform Gyrus (Anterior/Posterior division) • Primed vs. Unprimed: significant effect of condition x anterior/posterior at timepoints 4-6 (F(1,10)=5.704, p = 0.038; see Figure 4). • Primed vs. Exemplar: trend toward significance - condition x anterior/posterior interaction (F(1,10)=4.690, p = 0.052). • Posterior Fusiform Gyrus (see Figure 5) • Primed vs. Unprimed: differed significantly at timepoints 4-6 (t(10)=-2.523, p = 0.030) in left hemisphere and a trend towards significance in the right hemisphere (t(10)=-2.118, p = 0.060; see Figure 4) in the same time window. • Exemplar vs. Unprimed: no significant effects were observed in these regions. • Posterior Inferior Temporal Cortex (see Figure 5). • Exemplar vs. Unprimed: significant condition x hemisphere interaction at timepoints 4-6 • (F(1,8) = 25.466, p = 0.001; Figure 4). • Primed vs. Unprimed: no significant effects of repetition in this region • DISCUSSION AND CONCLUSIONS • REPETITION EFFECTS IN POSTERIOR FUSIFORM • As predicted, repetition suppression was observed in the fusiform gyrus in the left hemisphere. • This has been observed in previous masked word studies (e.g., Dehaene et al., 2001; 2004) and picture priming studies (e.g., Buckner et al., 1998; Koutstaal et al., 2001; Zago et al., 2005). • Furthermore, when dividing the fusiform gyrus into anterior and posterior portions, this effect is observed in the posterior fusiform bilaterally (trend for the right hemisphere). • This suggests early effects of repetition in the visual stream are due to visual similarity, especially since this effect is not observed for the different exemplar condition that differs in visual appearance. • Dehaene et al. (2004) observed a similar anterior/posterior sensitivity with masked word priming, which he suggested could be explained by fusiform neurons being tuned to progressively larger and more invariant units (from posterior to anterior). • The similarities in fusiform activation between Dehaene’s study with masked words and our study with masked pictures may suggest an overlap in picture and visual word form systems in the fusiform gyrus. • DIFFERENT EXEMPLAR EFFECT IN POSTERIOR ITG • In a distinct region of the posterior inferior temporal cortex an enhancement effect was observed in the left hemisphere for different exemplar compared to unprimed pictures, whereas repetition suppression was observed in the right hemisphere. The different exemplar pictures did not differ significantly in this region with primed pictures, although there was a trend towards significance (p=0.067). • This effect is the opposite observed by Koutstaal et al. (2001) with supraliminal pictures where different exemplar showed less repetition reduction in the right fusiform cortex, which could be due to the masked priming of pictures in the current experiment. The posterior inferior temporal cortex may be involved in a later process in the visual stream that takes into account the categorical nature of the stimuli. • POSSIBLE RELATIONSHIP TO ERP REPETITION EFFECTS Figure 4: ROI Analysis Anterior/Posterior Fusiform Gyrus Posterior ITG Figure 1: Typical Trials 300 msec 50 msec 10 msec 50 msec Repetition Condition (condition 1) 300 msec Forward Mask Prime Backward Mask Different Exemplar Condition (condition 2) Target Unrelated Condition (condition 3) Control Condition One possibility is that the masked repetition effects observed in fMRI correspond to the early repetition effects (100-300) observed in the occipital lobes in the analogous ERP study (figure is showing a occipital electrode site). It is less likely the fusiform repetition effect observed reflects the N400 since it has been established the N400 effect reflects semantic processing. Whereas, earlier effects are though to reflect lower level perceptual processing. FIG 2: Example slice orientation from 8-channel Siemens head coil Contact information: marianna.eddy@tufts.edu This research was supported by HD25889 and HD043251