Download

1 / 12
120 likes | 313 Views
Henry Lester June 2009. Engineering Ion Channels for Selective Neuronal Activation and Silencing. Neuronal Engineering with Cys-loop Receptor Channels. Goal: develop a general technique to selectively and reversibly silence or activate specific sets of neurons in vivo .

E N D
Henry Lester June 2009 Engineering Ion Channels for Selective Neuronal Activation and Silencing
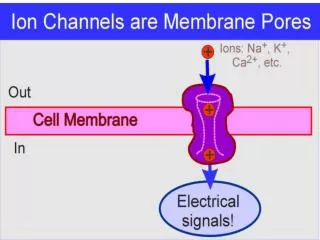
Neuronal Engineering with Cys-loop Receptor Channels Goal: develop a general technique to selectively and reversibly silence or activate specific sets of neurons in vivo. • Ideal approach would: • Have on- and off- kinetics on a time scale of minutes • Have simple activation (ie, via drug injected or in animal’s diet) • Avoid nonspecific effects in animal • Maintain target neurons healthy in an “off-state” for a few days without morphological/other changes • Silence or activate “diffuse” molecularly defined sets of neurons, not just spatially defined groups • The chosen channel • Cys-loop receptor (like nicotinic receptors) • Heteropentamer: α2β3 or α3β2 subunits. • This feature allows one to intersect two promoters, to enhance cellular specificity
The “channelohm” is 2% of the human genome, and many other organisms expand the repertoire Voltage (actually, ΔE ~107 V/m) External transmitter Internal transmitter Light Temperature Force/ stretch/ movement Blockers Binding region Switches Resistor Battery = Membrane region 1/r = 0.1 – 100 pS Nernst potential for Na+, K+, Cl-, Ca2+, H+ Colored by subunit (chain) Cytosolic region (incomplete) Invertebrate glutamate-gated Cl- channel . At this resolution, resembles nicotinic acetylcholine receptor
The drugs: “avermectins” • IVM: Lactone originally isolated from Streptomyces avermitilis • AVMs are used as antiparasitics in animals and humans (“River blindness” / Heartgard™) • IVM is probably an allosteric activator of GluCl channels • Also modulates GABA, 5HT3, P2X, and nicotinic channels, at much higher doses (IVM)
IVM-induced silencing in GluCl-expressing cultured rat hippocampal neurons 500 nm IVM 50 nm IVM 5 nm IVM
A B C D Optimized constructs optGluCla,b = “AVMR-Cl” • Binding site: • subunit unmutated; b Tyr182Phe (cation-π site) • suppresses endogenous glutamate sensitivity • M3-M4 intracellular loop: a YFP; b CFP • allows visualization • Coding region: codons adapted for mammalian expression • ~ 10-fold greater expression
AAV-2 constructs injected into mouse striatum; slice experiments Single neurons: correlation between IVM-induced conductance & AP silencing Lerchner et al, 2007 (collaboration with D. J. Anderson at Caltech)
Plans to extend the AVMR system Transfer AVM sensitivity to mammalian glycine receptor no immune response Tighter AVM binding increased AVM sensitivity M2 mutations increased AVM sensitivity • Na+-permeable • selective neuronal activation • Ca2+-permeable • manipulate signal transduction Increased single-channel current increased AVM sensitivity Optimize ER exit and trafficking → increased surface expression M3-M4 loop
Very slow (several hr) AVM reversibility is puzzling GluCl- heteromer GluCl- homomer No potentiation GluCl- homomer (Etter et al., JBC 1996)
1mM Glu 1mM IVM Cys-loop 89 loop Location of the AVM binding site is unknown Likely distinct from the glutamate binding site Covalent binding interaction? Within the cavity of the TMD? At the ECD-TMD interface? (where other anesthetics are bind) Yoav Paas, BIU McCammon Lab, UCSD Radioligand binding experiments with [3H]-IVM on C. elegans membrane preps IVM binding sites exhibit high affinity binding (KD = 0.11 nM) IVM does dissociate from its receptor, with a rate constant of 0.005-0.006/min (Cully & Paress, 1991)
The first AVMR-Na GluCl WT + WT Muscle nAChR ND98 (10 nM IVM) 0.5 ND98 GluCl P(-2’)/A(-1’)E + WT Still too small Still too large (200 nM IVM)
Many AVMRs remain in intracellular compartments, but are chaperoned by IVM (GluClαYFP)GluClβ 24 h incubation (control solution) The intensity ratio, peripheral/whole cell, is 0.86 ± 0.07 in control and 1.51 ± 0.10 in IVM-treated cells (SEM (1 μM IVM) Confocal TIRF