Download

1 / 44
450 likes | 673 Views
Gerhard Krauss Biochemistry of Signal Transduction and Regulation(3rd Edition) ISBN: 3-527-30591-2. 授課老師 : 褚俊傑副教授 ( 生物科技系暨研究所 ) 聯絡電話 : 0986-581835 電子信箱 : jjchuu@mail.stut.edu.tw. Signaling by Nuclear Receptors. Nuclear Receptor Signaling Pathways. Outline.

E N D
Gerhard Krauss Biochemistry of Signal Transduction and Regulation(3rd Edition) ISBN: 3-527-30591-2 授課老師: 褚俊傑副教授 (生物科技系暨研究所) 聯絡電話: 0986-581835 電子信箱: jjchuu@mail.stut.edu.tw Signaling by Nuclear Receptors
Outline • 4.1 Ligands of Nuclear Receptors • 4.2 Principles of Signaling by Nuclear Receptors • 4.3 Classification and Structure of Nuclear Receptors • 4.4 Mechanisms of Transcriptional Regulation by Nuclear Receptors • 4.5 Regulation and Variability of Signaling by Nuclear Receptors • 4.6 The Signaling Pathway of the Steroid Hormone Receptors • 4.7 Signaling by Retinoids, Vitamin D3, and the T3-Hormone
4.1 Ligands of Nuclear Receptors • The naturally occurring ligands of nuclear receptors are lipophilic hormones, among which the steroid hormones, the thyroid hormone T3, and derivatives of vitamin A and D have long been known as central regulators. • These hormones play a significant role in metabolic regulation, organ function, and development and differentiation processes. Following formation and secretion in specific tissues, the endocrine organs, the hormones are distributed in the organism via the circulation and enter cells passively by diffusion.
4.1 Ligands of Nuclear Receptors In recent years it has been recognized that intracellularly formed lipophilic metabolites can also serve as ligands for nuclear receptors and can regulate gene expression through their binding to nuclear receptors. These compounds include prostaglandins, leukotrienes, fatty acids, cholesterol derivatives, bile acids, and even benzoates. The most important natural ligands of the nuclear receptors are shown in Fig. 4.1; the cognate receptors and their DNA elements are summarized in Table 4.1.
Tab. 4.1 Ligands and structure of HREs of selected nuclear receptors from mammals. Abbreviations: IR: “inverted repeat”; DR: “direct repeat”; ER: “everted repeat”; NR: “no repeat”. Numbers of HREs give the number of pairs separating the half-sites. R: purine. Y: pyrimidine. NGFI: Nerve growth factor induced receptor; COUP-TF: CHicken ovalbumin upstream promoter transcription factor; ROR: Retinoic acid related orphan receptor. a, b und c are receptor subtypes coded by distinct genes.
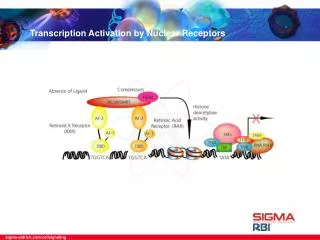
4.2 Principles of Signaling by Nuclear Receptors • Signal transduction by nuclear receptors is shown schematically in Fig. 4.2. The nuclear receptors are localized in the cytosol and/or nucleus. Many of the natural ligands of nuclear receptors are lipophilic hormones that enter the cell in a passive manner or by active transport mechanisms. • A transmittance of the signal at the cell membrane is not necessary, as is the case in the signaling via transmembrane receptors. Once inside the cell, the hormone ligand binds the cognate receptor which is localized in the cytosol and/or in the nucleus. The hormone binding activates the transcription regulation function of the receptor. • In the case of the cytosolic receptors, the hormone binding induces translocation into the nucleus where the hormone-receptor complex binds a cognate DNA element termed hormone-responsive element, HRE, and alters the transcription of the target gene.
Fig. 4.2 The principle of signal transduction by nuclear receptors. Nuclear receptors are ligand controlled transcritpion factors that bind cognate DNA sequences, or hormone responsive elements (HRE). The hormone acts as a regulating ligand. Most nuclear receptors bind their cognate HRE, which tend to be symmetrically organized, as homo- or heterodimers. The DNA bound, activated receptor stimulates transcription initiation via direct or indirect protein-protein interactions with chromatin and the transcription initiation complex. The arrows demonstrate the different possible configurations of the HRE (see also Section 4.3.1). H: hormone; Hsp: heat shock protein.
4.3 Classification and Structure of Nuclear Receptors • 4.3.1 DNA-Binding Elements of Nuclear Receptors, HREs • 4.3.2 The DNA-Binding Domain of Nuclear Receptors • 4.3.3 HRE Recognition and Structure of the HRE-Receptor Complex • 4.3.4 Ligand-binding Domains • 4.3.5 Transactivating Elements of the Nuclear Receptors
4.3 Classification and Structure of Nuclear Receptors • The nuclear receptors display a high degree of homology at the level of amino acid sequence, which indicates that they operate with similar functional principles. • For some receptors the cognate hormone and their function in the cell remain unknown. Such “orphan receptors” are usually identified by sequence homology and with the help of screening techniques employing DNA probes based on known receptors. Possibly some of the orphan receptors do not have natural ligands and function in a ligand-independent manner. • At the level of the primary structure the steroid hormone receptors can be divided into five different domains(Fig. 4.3), each with specific functions.
Fig. 4.3 Domain structure of the nuclear receptors. Functional domains of nucear receptors are portrayed in a one dimensional, linear fashion.
4.3.1 DNA-Binding Elements of Nuclear Receptors, HREs • The steroid hormone receptors are sequence-specific DNA-binding proteins whose cognate DNA elements are termed “hormone-responsive elements” (HREs). • The identity of an HRE is determined by the sequence, polarity, and distance of the hexamers. Mutation and duplication of an ancestral recognition sequence have allowed the creation of many and various DNA elements during the course ofevolution, whose sequence, polarity and distance is characteristic for a given hormone receptor or receptor pair.
4.3.1 DNA-Binding Elements of Nuclear Receptors, HREs The half-site of an HRE can be arranged as a palindrome, an inverted palindrome or a direct repeat. For a given receptor, optimal spacings of the half-sites exist, and the number of base pairs between the half-sites is another charactistic feature of a HRE. Figure 4.4 illustrates, on the example of the HREs for the RXR heterodimer, the various configurations of an HRE.
Fig. 4.4 HRE structure of the RXR heterodimer. Shown is the consensus sequence of the HREs of the RXR heterodimers (see Fig. 4.7) and the different possible arrangements of the hexameric half-site sequences. The hexamers can be arranged palindromically as inverted repeats (a), as everted repeats (b), or as direct repeats (c). n indicates the number of base pairs that lie between the two hexamers. RXR: receptor for 9-cis-retinoic acid; RAR: receptor for all-trans retinoic acid; T3R: receptor for the T3 hormone; PPAR peroxisome proliferator activated receptor; VDR: receptor for vitamin D3.
4.3.1 DNA-Binding Elements of Nuclear Receptors, HREs • Based on the subunit structure of DNA-bound receptors and on the structure of the HREs, four classes of nuclear receptors can be distinguished (Fig. 4.5). • Dimers of the Steroid Hormone Receptors The HREs of the steroid hormone receptors possess a palindromic structure, comparable to the DNA-binding elements of procaryotic repressors(see Fig. 4.5a). • Heterodimers Containing RXR The DNA-binding elements of the nuclear receptors for all-trans retinoic acid, for 9-cis retinoic acid, for the T3 hormone and for the vitamin D3 hormone usually exhibit a direct repeat of the recognition sequence, resulting in the formation of heterodimers on the DNA(Fig. 4.5b).
4.3.1 DNA-Binding Elements of Nuclear Receptors, HREs Dimeric Orphan Receptors The “orphan receptors” derive their name from the fact that the cognate hormones for these receptors were originally unknown or little understood. Orphan receptors can bind as homodimers to recognition sequences arranged as direct repeats(Fig. 4.5c). Monomeric “Orphan Receptors” Orphan receptors are also known (e.g., Retinoic Z receptor, RZR; NGF-induced clone B, NGFI-B) which bind as monomers to asymmetric recognition sequences (Fig. 4.5d).
Fig. 4.5 Oligomeric structure of nuclear receptors and structure of the HREs. The nuclear receptors can be subdivided into four groups based on strucutres of the receptors and HREs. Shown above are some representative examples. a) binding of a homodimeric receptor to a two-fold symmetric palindromic DNA element, GR: gluccocorticoid receptor. b) binding of a heterodimeric receptor to a DNA element with direct repeats of the recognition sequence, whereby the 5’ side of the HRE is occupied by the receptor for 9-cis retinoic acid (RXR). RAR: receptor for all-trans retinoic acid, T3R: receptor for the T3 hormone; PPAR: peroxisome proliferating activated receptor; VDR: receptor for vitamin D3. c) binding of RXR as a homodimer to an HRE with direct repeat of the recognition sequence. d) binding of a monomeric receptor to an asymmetric recognition sequence, NGFI-B: nerve growth factor induced receptor, is involved in the regulation of enzymes of steroid biosynthesis.
4.3.2 The DNA-Binding Domain of Nuclear Receptors • The binding of receptors to their cognate HRE occurs via a DNA-binding domain, which are largely independent folding domains. Within the family of nuclear receptors, the DNA-binding domain is the most conserved structural element and is located in region C of the primary structure (see Fig. 4.6). • The DNA-binding domain possesses structural elements that mediate the specific recognition of the HRE, as well as for the dimerization of the receptor on the HRE. The core of the DNA-binding domain includes a span of 70–80 amino acids, in which all information for the specific recognition of the cognate half-site is contained.
Fig. 4.6 Functional domains, DNA binding and HRE structure of nuclear receptors on the example of the glucocorticoid receptor, GR. a) domain structure of GR. AF1, AF2: domains that mediate the stimulation of the transcription. b) schematic representation of the two Zn2+-Cys4 binding motifs of the DNA binding domains. c) Complex formation between the dimeric DNA binding domains of GR and the HRE. The black spheres represent Zn2+ ions. d) Consensus sequence and configuration of the HRE elements of GR.
4.3.3 HRE Recognition and Structure of the HRE-Receptor Complex • For direct repeat HREs, the spacing of the two half-sites is often the decisive, if not the only, element based on which the receptor (homodimer or heterodimer) recognizes its own HRE and discriminates against related HREs. • The solution of the structures of heterodimer-DNA complexes has shown how these receptors can distinguish between highly related HREs(Fig.4.7). As an example, the structure of a DNA-bound receptor-heterodimer composed of the DNA-binding domain of RXR and the T3-receptor is given in Fig. 4.7.
Fig. 4.7 Structure of the RXR-T3R heterodimer in complex with DNA. Illustrated is a complex between the DNA binding domain of the RXR-T3R heterodimer and an HRE with direct repeats of the sequence AGGTCA separated by 4 base pairs. The two receptor subunits contact the hexameric sequences with a recognition helix in a manner very similar to that of the gluccocorticoid receptor (see Fig. 4.6). The Zn atoms are drawn as green spheres. The figure illustrates the polarity of the binding of the two subunits. The interaction between the two subunits is mediated mainly via an extension of the C-terminal DNA binding domain (bottom) of the T3R. A greater or smaller distance between the two hexamers of the HRE would act contrary to the interaction between the two subunits as shown.
4.3.4 Ligand-binding Domains • The region E with the ligand-binding domain harbors three important functions: – homo- and heterodimerization – binding of ligand, both agonists and antagonists – transactivation and transrepression: binding of coactivators and corepressors. • Dimerization A contribution to the dimerization of the receptors – in addition to that from the DNA binding domain – is provided by a dimerization element in the ligand-binding domain. The structure of the ligand-binding domain of RXR without bound hormone shows a homodimer with a symmetric dimerization surface, formed essentially from two antiparallel a-helices.
4.3.4 Ligand-binding Domains • Ligand Binding The crystal structures of the ligand-binding domains of several nuclear receptors have been resolved demonstrating a similar overall structure. Fig. 4.8a shows the receptor for alltrans retinoic acid, RAR, with is bound ligand. The ligand-binding domain is formed from 12 a-helices numbered from H1 to H12. In the bottom half of the structure, a ligand-binding pocket is found, which accommodates the ligand. The pocket is mainly hydrophobic and of variable size for different receptors. As illustrated in Fig. 4.8b for the binding of RAR bound to all-trans-retinoic acid, contacts between the ligand and the pocket can be very extensive and include many hydrophobic contacts as well as hydrogen bonds to the polar parts of the ligand. Fig. 4.8 a) Ribbon diagram of RAR bound to all trans retinoic acid. b) Schematic diagram of retinoic acid binding site. H-bonds and ionic interactions fix the carboxylate of the ligand whose nonpolar parts are deeply buried in a hydrophobic pocket.
4.3.4 Ligand-binding Domains Fig. 4.9 Structural changes in the ligand binding domain of RXR on binding of 9-cis retinoic acid. The models of domain E of apo-RXRα and the binary complex of RXRα and 9-cis retinoic acid were superimposed. Domain E of apo-RXRα is depicted in green and yellow, and domain E of the binary complex in blue and red. The grey arrows indicate the structural rearrangements of helices 11, 12, and the N-terminus of helix 3. Ligand Binding The H12 helix is amphipathic, possessing a hydrophobic and a hydrophilic face. In the unliganded RXR, helix 12 projects away from the body of the ligand-binding domain. In the liganded structure, the helix reorients (Fig. 4.9). As illustrated in Fig. 4.10, the structure of a ligand-binding domain with an antagonist bound can provide a rational basis to explain the antagonistic function of a ligand.
Fig. 4.10 Agonist vs antagonist binding to estrogen receptor, ER a) Ribbon diagram of ER bound to the agonist estradiol (black). b) Ribbon diagram of ER bound to the antagonist raloxifene (black). c) schematic drawing of estradiol-binding site. d) schematic diagram of raloxifene-binding site.
4.4 Mechanisms of Transcriptional Regulation by Nuclear Receptors • Most of the functions of nuclear receptors can be described in terms of activation and repression of transcription. Although attention has been focussed primarily on the transcription activation mediated by “positive” HREs, it is increasingly recognized that nuclear receptors can also repress transcription in a ligand-dependent manner.
4.4 Mechanisms of Transcriptional Regulation by Nuclear Receptors So-called “negative HREs” have been identified that bind the receptor and mediate negative regulation by the ligand. These elements have been identified for glucocorticoid receptors and for the T3 receptor. In addition to ligand-dependent activation and repression, a subset of nuclear receptors repress basal transcription in the absence of ligand when bound to a positive HRE, thereby silencing the target gene(Fig. 4.11).
Fig. 4.11 Model of repression and activation of T3R. In the absence of the T3 hormone, a heterodimeric RXR-T3R receptor is bound at the T3-responsive element,TRE, establishing a basal repressed state. The repressed state is maintained by recruitment of corepressor complexes containing histone deacetylase activity. X refers to potential unidentified cofactors (possibly chromatin remodeling complexes or SRBs) which help to keep the promotor-bound basal transcription apparatus in the inactive state. In the presence of T3 hormone, the corepressors are removed and coactivators (e.g. the SRC/p160 complex) bind to the receptor heterodimer. The histone acetylase activity of the associated proteins helps to induce a transcription competent state of the nucleosomes. Furthermore, another coactivator complex, DRIP/TRAP, is recruited by to the promotor which is thought to stabilize binding of the RNA polymerase holoenzyme.
4.4 Mechanisms of Transcriptional Regulation by Nuclear Receptors • Coactivators of Nuclear Receptors A series of proteins or protein complexes with coactivator function for nuclear receptors has been identified that specifically interact with the activated, liganded receptor. The most abundant ones are now included in the p160 family of coactivators with the steroid receptor coactivator 1, SRC-1, as a well-characterized member. Another group of coactivators is present as multiprotein complexes like the TRAP complex (TRAP, thyroid hormone receptor activating protein). A common receptor interaction motif LXXLL is found on these coactivators which mediates at least part of the interaction with the AF2 domain of the receptor.
4.4 Mechanisms of Transcriptional Regulation by Nuclear Receptors • Corepressors of Nuclear Receptors Unliganded T3 receptor (T3R) and retinoic acid receptor (RAR) can repress transcription in the absence of ligand. This transcriptional repression is linked to the binding of proteins with corepressor activity. Examples of such corepressors are the nuclear corepressor NcoR and the “silencing mediator for retinoic and thyroid hormone receptors”, SMRT. Both proteins interact with the ligand-binding domain and appear to be released from the receptor upon ligand binding. Possibly alternative conformations of the AF-2 region serve to trigger this release.
4.5 Regulation and Variability of Signaling by Nuclear Receptors • Signaling by nuclear receptors is regulated at various levels (Fig. 4.12). The following are important regulatory attack points: • Regulation at the level of ligand concentration • Crosstalk: regulation by phosphorylation • Interaction with other transcriptional activators • Regulation by ubiquitination
Fig. 4.12 Functions of nuclear receptor domains. The domains A/B, C, E and F of the nuclear receptors are involved in multiple protein-protein interactions and are subject to regulatory modifications as indicated. Most important are the corepressor and coactivator complexes that direct histone deacetlyase (HDAC) and histone acteylase (HAT) activities, respectively, to the nuclear receptor regulated promotor region. TRAP: thyroid hormone receptor activating protein.
4.5 Regulation and Variability of Signaling by Nuclear Receptors * Regulation at the level of ligand concentration A main determinant of nuclear receptor signaling is the concentration of the ligand available for binding. The ligand concentration can be regulated in many ways (for details see textbooks on hormone action): – synthesis and degradation – modification – secretion, transport and storage – feedback regulation via the circulating hormone concentration
4.5 Regulation and Variability of Signaling by Nuclear Receptors * Crosstalk: regulation by phosphorylation The phosphorylation of nuclear receptors on Ser/Thr residues is a key mechanism for the coupling of nuclear receptor signaling to other signaling pathways of the cell. Many nuclear receptors are isolated as phosphoproteins, and their phosphorylation provides a means for ligand-independent activation and regulation.
4.5 Regulation and Variability of Signaling by Nuclear Receptors * Interaction with other transcriptional activators Nuclear receptors can also modulate gene expression by interference with the activity of other transcriptional activators. The ER, T3R, RAR and GR proteins have, e.g., been shown to act as transrepressors of the transcription factor AP-1, which is a heterodimer composed of c-Jun and c-Fos proteins. Reciprocally, AP-1 can inhibit transactivation by these receptors. * Regulation by ubiquitination As for other central regulators, the level of nuclear receptors is modulated by ubiquitin-mediated degradation. A ligand-dependent ubiqutination and subsequent proteasomal degradation has been described for ERa, PR, VDR, T3R and RARa, allowing for a down-regulation of signaling by these receptors.
4.6 The Signaling Pathway of the Steroid Hormone Receptors • Based on the receptor activation mechanism, the nuclear receptors may be divided into two basic groups. In the first group (those including most of the steroid hormone receptors), the receptors can be localized in the nucleus or in the cytoplasm. • Signal transduction by steroid hormones is distinguished by the fact that the receptors can be found either in the cytoplasm or in the nucleus. The steroid hormone receptor receives the hormonal signal in the cytosol, becomes activated by hormone binding, at which point it enters the nucleus to regulate the transcription initiation of cognate genes. • Figure 4.13 shows the most important steps in the signal transduction by steroid hormones.
Fig. 4.13 Principle of signal transduction by steroid hormone receptors. The steroid hormone receptors in the cytosol are found in the form of an inactive complex with the heat shock proteins Hsp90 and Hsp56 and with protein p23. The binding of the hormone activates the receptor so that it can be transported into the nucleus where it can bind to its cognate HRE. It remains unclear in which form the receptor is transported into the nucleus, and to which extent the associated proteins are involved in the transport. One mechanism of activation of transcription initiation involves mediation by the proteins RIP-140 and Sug1.
4.7 Signaling by Retinoids, Vitamin D3, and the T3-Hormone • 4.7.1 Structure of the HREs of RXR Heterodimers • 4.7.2 Complexity of the Interaction between HRE, Receptor and Hormone
4.7 Signaling by Retinoids, Vitamin D3, and the T3-Hormone • In contrast to signal transduction by the steroid hormone receptors, there are multiple pathways by which the ligands of this group are made available for receptor activation (Fig. 4.14): * The hormone ligands can be secreted in the classical endocrinological pathway and transported to the target cell where they bind the receptor. * The active hormones can be formed intracellularly from inactive precursors. The inactive precursor is transported through the bloodstream to the target cell where it is enzymatically converted to the active hormone. An example of this pathway is that of 9-cis retinoic acid, which is synthesized from the alcohol of vitamin A (vitamin A1; retinol).
Fig. 4.14 Principle of signal transduction by RXR heterodimers. The activated hormone can be made available to the RXR heterodimer in three different ways. a) The hormone (e.g. T3 hormone) is synthesized in endocrinal tissue and reaches the DNA bound RXR heterodimer in the nucleus via passive transport. b) The active hormone is formed in the cytosol from an inactive apo-hormone (as for, e.g. 9-cis-retinoic acid). c) The hormone is synthesized intracellularly. In all three cases, the binding of the hormone-RXR-heterodimeric complex is the signal that induces transcription activation of the downstream genes.
4.7.1 Structure of the HREs of RXR Heterodimers • The following points were identified as important for the recognition and discrimination of a particular HRE: • In the case of identical hexamer sequences of an HRE, the spacing between the hexamers is a specificity-determining element (n-rule). The spacing can be between 1 and 6 bp. Grounds for the discrimination based on spacing is the structure of the receptor dimer. A given receptor demands a particular spacing of the hexamers in the HRE because of steric requirements. • The receptor for 9-cis retinoic acid (RXR) usually occupies the 5’ position in the heterodimer. The RXR serves as a quasi vehicle to bring other receptor monomers to the 3’ half-site of the HRE.
4.7.2 Complexity of the Interaction between HRE, Receptor and Hormone • There is an altogether complex interaction between HRE, receptor and hormone in the group of receptors for retinoids, vitamin D3 and the T3-hormone. The complexity is determined by the following factors: * Formation of the Homo- or Heterodimer * Multiplicity of the HREs * Multiplicity of the Receptors * Binding and Activation via Hormones