Download
1 / 36
370 likes | 524 Views
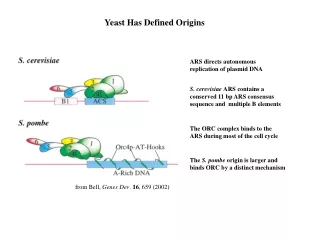
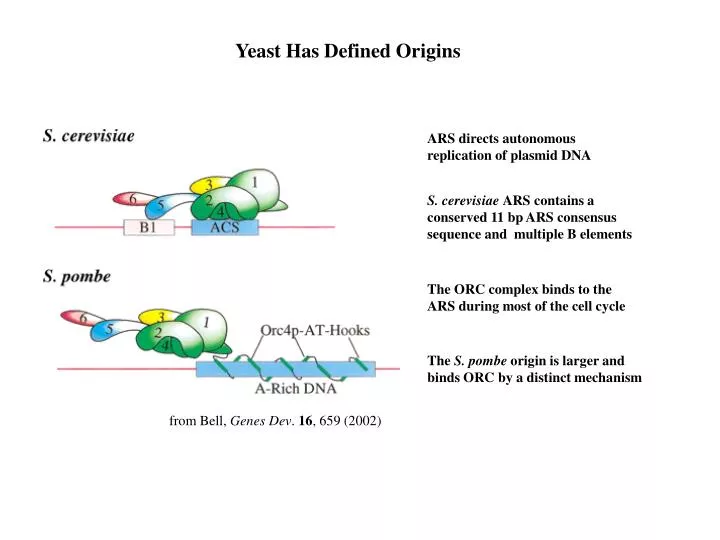
Yeast Has Defined Origins. ARS directs autonomous replication of plasmid DNA. S. cerevisiae ARS contains a conserved 11 bp ARS consensus sequence and multiple B elements. The ORC complex binds to the ARS during most of the cell cycle. The S. pombe origin is larger and

E N D
Yeast Has Defined Origins ARS directs autonomous replication of plasmid DNA S. cerevisiae ARS contains a conserved 11 bp ARS consensus sequence and multiple B elements The ORC complex binds to the ARS during most of the cell cycle The S. pombe origin is larger and binds ORC by a distinct mechanism from Bell, Genes Dev. 16, 659 (2002)
Replication Origins in Metazoans DNA replication initiates from distinct confined sites or extended initiation zones The potential to initiate is modulated by sequence, supercoiling, transcription, or epigenetic modifications Initiation can influence initiation at an adjacent site from Aladjem, Nature Rev.Genet.8, 588 (2007)
Some Features of Eukaryotic Replication Origins from Méchali, Nature Rev.Mol.Cell.Biol. 11, 728 (2010) Certain characteristics are common at metazoan replication origins but are not present at all origins Different modules contribute to the selection of a given origin
Different Classes of Replication Origins in Metazoans Only a small subset of origins are active during a given cell cycle Constitutive origins are used all the time and are relatively rare Flexible origins are used to a different extent in different cells and follow the Jesuit Model “Many are called but few are chosen” Inactive or dormant origins are only used during replication stress or during certain cellular programs from Méchali, Nature Rev.Mol.Cell.Biol. 11, 728 (2010)
Chromatin Structure Influences ORC Binding Chromatin remodelling complexes can facilitate HAT binding preRC proteins can be modified by HATs from Méchali, Nature Rev.Mol.Cell.Biol. 11, 728 (2010)
Influence of Distal Elements on Initiation Deletion of DHFR promoter allows initiation to occur within the gene Truncation of the DHFR gene confines initiation to the far end of the locus Deletion of the b-globin LCR prevents initiation within the locus Deletion of the CNS1 sequence in the Th2 cluster do not initiate within the IL13 gene from Aladjem, Nature Rev.Genet.8, 588 (2007)
The Formation of the preRC Mcm2-7 is loaded as a double hexamer by ORC, Cdc6 and Cdt1 Sld3 and Cdc45 bind weakly to Mcm2-7 Mcm2-7 helicase is inactive until S phase from Labib, Genes Dev. 24, 1208 (2010)
Origins Are Activated at Different Times preRCs are formed during G1 on origins Heterochromatic regions replicate later than euchromatic regions from Méchali, Nature Rev.Mol.Cell.Biol. 11, 728 (2010)
The Replicative Helicase Mcm2-7, Cdc45, and GINS (CMG complex) form the replicative helicase from Moyer et al., Proc.Nat.Acad.Sci.USA 103, 10236 (2006)
Assembly of the Replicative Helicase preRC is formed during G1 by recruitment of Mcm2-7 Phosphorylation of MCM proteins by DDK recruits GINS and stabilizes Cdc45 association from Sheu and Stillman, Mol.Cell 24, 101 (2006)
Helicase Loading and Activation in DNA Replication DnaA and ORC are structural homologs Replication competence is conferred by Mcm2-7 loading and is prevented by inhibition of pre-RC proteins CDKs prevent Mcm2-7 loading and are required for helicase activation from Remus and Diffley, Curr.Opin.Cell Biol. 21, 771 (2009)
Activation of Helicase Requires Phosphorylation of Sld2 and Sld3 G1 CDKs allow Dbf4 to accumulate DDK phosphorylates Mcm2-7 and promotes Cdc45 association CDK phosphorylates Sld2 and Sld3 and promotes association with Dpb11 from Botchan, Nature445, 272 (2007) 11-3-2 promotes helicase activation
Initiation of Chromosome Replication DDK phosphorylates Mcm proteins CDK phosphorylates Sld2 and Sld3 to interact with Dpb11 GINS and Pol e are recruited to form the RPC (replisome progression complex) Activation of the helicase allows priming by Pol a Pol e extends the leading strand and Pol d extends each Okazaki fragment from Labib, Genes Dev. 24, 1208 (2010)
Replication Origins are Licensed in Late M and G1 Origins are licensed by Mcm2-7 binding to form part of the pre-RC Mcm2-7 is displaced as DNA replication is initiated Licensing is turned off at late G1 by CDKs and/or geminin from Blow and Dutta, Nature Rev.Mol.Cell Biol.6, 476 (2005)
Control of Licensing Differs in Yeasts and Metazoans CDK activity prevents licensing in yeast Geminin activation downregulates Cdt1 in metazoans from Blow and Dutta, Nature Rev.Mol.Cell Biol.6, 476 (2005)
Telomeres are Specialized Structures at the Ends of Chromosomes Telomeres contain multiple copies of short repeated sequences and contain a 3’-G-rich overhang Telomeres are bound by proteins which protect the telomeric ends initiate heterochromatin formation and facilitate progression of the replication fork from Gilson and Geli, Nature Rev.Mol.Cell Biol. 8, 825 (2007)
Functions of Telomeres Telomeres protect chromosome ends from being processed as a ds break End-protection relies on telomere-specific DNA conformation, chromatin organization and DNA binding proteins from Gilson and Geli, Nature Rev.Mol.Cell Biol. 8, 825 (2007)
The End Replication Problem Leading strand is synthesized to the end of the chromosome Lagging strand utilizes RNA primers which are removed The lagging strand is shortened at each cell division from Lodish et al., Molecular Cell Biology, 6th ed. Fig 6-49
Solutions to the End Replication Problem 3’-terminus is extended using the reverse transcriptase activity of telomerase Dipteran insects use retrotransposition with the 3’-end of the chromosome as a primer Kluyveromyces lactis uses a rolling circle mechanism in which the 3’-end is extended on an extrachromosomal template Telomerase-deficient yeast use a recombination- dependent replication pathway in which one telomere uses another telomere as a template Formation of T-loops using terminal repeats allow extension of invaded 3’-ends from de Lange, Nature Rev.Mol.Cell Biol. 5, 323 (2004)
Telomerase Extends the ss 3’-Terminus Telomerase-associated RNA base pairs to 3’-end of lagging strand template Telomerase catalyzes reverse transcription to a specific site 3’-end of DNA dissociates and base pairs to a more 3’-region of telomerase RNA Successive reverse transcription, dissociation, and reannealing extends the 3’-end of lagging strand template New Okazaki fragments are synthesized using the extended template from Lodish et al., Molecular Cell Biology, 6th ed. Fig 6-49
The Action of Telomerase Solves the Replication Problem New Okazaki fragments are synthesized using the extended template from Alberts et al., Molecular Biology of the Cell, 4th ed. Fig 5-43
Shelterin Specifically Associates with Telomeres Shelterin subunits specifically recognize telomeric repeats Shelterin allows cells to distinguish telomeres from sites of DNA damage from de Lange, Genes Dev. 19, 2100 (2005)
Telomere Termini Contain a 3’-Overhang A nuclease processes the 5’-end POT1 controls the specificity of the 5’-end from de Lange, Genes Dev. 19, 2100 (2005)
Structure of Human Telomeres Telomeres consist of numerous short dsDNA repeats and a 3’-ssDNA overhang The G-tail is sequestered in the T-loop Shelterin is a protein complex that binds to telomeres TRF2 inhibits ATM-dependent DNA damage response Shelterin components block telomerase activity from O’Sullivan and Karlseder, Nature Rev.Mol.Cell Biol. 11, 171 (2010)
Telomerase Action is Restricted to a Subset of Ends Telomere length is regulated by shelterin Increased levels of shelterin inhibits telomerase action Telomerase is inhibited by increased amounts of POT1 Elongation of shortened telomeres depends on the recruitment of the Est1 subunit of telomerase by Cdc13 end-binding protein from Bertuch and Lundblad, Curr.Opin.Cell Biol. 18, 247 (2006)
Dysfunctional Telomeres Induce the DNA Damage Response Shelterin may contain an ATM inhibitor Telomere damage activates ATM DNA damage response proteins accumulate at unprotected telomeres ATM activates p53 and leads to cell cycle arrest or apoptosis from de Lange, Genes Dev. 19, 2100 (2005)
Loss of Functional Telomeres Results in Genetic Instability Dysfunctional telomeres activate DSB repair by NHEJ Fused chromosomes result in chromatid break and genome instability from O’Sullivan and Karlseder, Nature Rev.Mol.Cell Biol. 11, 171 (2010)
Loss of Telomeres Limits the Number of Rounds of Cell Division Stem cells and germ cells contain telomerase which maintains telomere size Somatic cells have low levels of telomerase and have shorter telomeres Loss of telomeres triggers chromosome instability or apoptosis Cancer cells contain telomerase and have longer telomeres from Lodish et al., Molecular Cell Biology, 6th ed. Fig 25-31
Telomerase-based Cancer Therapy Telomerase is widely expressed in cancers 80-90% of tumors are telomerase-positive Strategies include Direct telomerase inhibition Telomerase immunotherapy
Endogenous DNA Damage from Marnett and Plastaras, Trends Genet. 17, 214 (2001)
Biological Molecules are Labile RNA is susceptible to hydrolysis Reduction of ribose to deoxyribose gives DNA greater stability N-glycosyl bond of DNA is more labile DNA damage occurs from normal cellular operations and random interactions with the environment
Spontaneous Changes that Alter DNA Structure deamination oxidation depurination from Alberts et al., Molecular Biology of the Cell,4th ed., Fig 5-46
Hydrolysis of the N-glycosyl Bond of DNA from Alberts et al., Molecular Biology of the Cell,4th ed., Fig 5-47 Spontaneous depurination results in loss of 10,000 bases/cell/day Causes formation of an AP site – not mutagenic
Deamination of Cytosine to Uracil from Alberts et al., Molecular Biology of the Cell,4th ed., Fig 5-47 Cytosine is deaminated to uracil at a rate of 100-500/cell/day Uracil is excised by uracil-DNA-glycosylase to form AP site
5-Methyl Cytosine Deamination is Highly Mutagenic Deamination of 5-methyl cytosine to T occurs rapidly - base pairs with A 5-me-C is a target for spontaneous mutations from Alberts et al., Molecular Biology of the Cell,4th ed., Fig 5-52
Deamination of A and G Occur Less Frequently A is deaminated to HX – base pairs with C G is deaminated to X – base pairs with C from Alberts et al., Molecular Biology of the Cell,4th ed., Fig 5-52