Download
1 / 23
230 likes | 356 Views
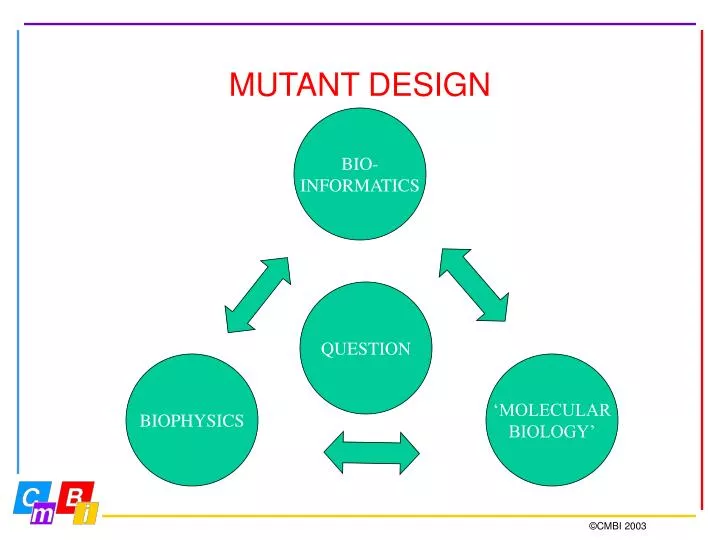
BIO- INFORMATICS. QUESTION. BIOPHYSICS. ‘MOLECULAR BIOLOGY’. MUTANT DESIGN. BIO- INFORMATICS. QUESTION. BIOPHYSICS. ‘MOLECULAR BIOLOGY’. MUTANT DESIGN. Abstract Protein folding, structure, stability Applied Process optimization. MUTANT DESIGN.

E N D
BIO- INFORMATICS QUESTION BIOPHYSICS ‘MOLECULAR BIOLOGY’ MUTANT DESIGN ©CMBI 2003
BIO- INFORMATICS QUESTION BIOPHYSICS ‘MOLECULAR BIOLOGY’ MUTANT DESIGN Abstract Protein folding, structure, stability Applied Process optimization ©CMBI 2003
MUTANT DESIGN • Three strong warnings and disclaimers: • I know nothing about MAKING mutants • Most times ‘evolutionary’ (that is grant-writing terminology for smart trial-and-error) beat design approaches. • Mutants are not always the best way to answer questions. Often good old-fashioned protein chemistry, spectroscopy, or even literature searches get you the answer more quickly. ©CMBI 2003
WHY MUTATIONS • Understand protein folding, structure, stability (against many different things); • Atomic model validation (homology models, drug binding), or abstract model validation (functional hypotheses); • Disrupting interactions, or make them permanent; • Protein activity is very hard to engineer; • Support for structure determination, e.g. Selenomethionine for SAD or MAD, Cysteine for heavy-metal binding, solubility for NMR; introduce fluorophore; • Humanization (normally more than just mutations); • Delete, or sometimes add post-translational modifications; • Purification tags, e.g. his-tag, flag-tag (not really mutations); • Temperature sensitive mutants; • Alanine or cysteine scan, or variants thereof; • ‘Mutate away’ metal binding sites; Many mutations belong in more than one category….. ©CMBI 2003
PROTEIN STRUCTURE helix strand turn Alanine 1.42 0.83 0.66 Arginine 0.98 0.93 0.95 Aspartic Acid 1.01 0.54 1.46 Asparagine 0.67 0.89 1.56 Cysteine 0.70 1.19 1.19 Glutamic Acid 1.39 1.17 0.74 Glutamine 1.11 1.10 0.98 Glycine 0.57 0.75 1.56 Histidine 1.00 0.87 0.95 Isoleucine 1.08 1.60 0.47 Leucine 1.411.30 0.59 Lysine 1.14 0.74 1.01 Methionine 1.45 1.05 0.60 Phenylalanine 1.131.38 0.60 Proline 0.57 0.55 1.52 Serine 0.77 0.75 1.43 Threonine 0.83 1.19 0.96 Tryptophan 1.08 1.37 0.96 Tyrosine 0.69 1.47 1.14 Valine 1.06 1.70 0.50 Abstract Applied ©CMBI 2003
PROTEIN STABILITY ΔG = ΔH - TΔS ΔG = -RT ln(K) K = [Folded] / [Unfolded] So, you can interfere either with the folded, or with the unfolded form. Choosing between ΔH and ΔS will be much more difficult, because ΔG is a property of the complete system, including H2O…. ©CMBI 2003
PROTEIN STABILITY Hydrophobic packing Helix capping Loop transplants ©CMBI 2003
PROTEIN STABILITY A whole series of tricks can be applied: Gly -> Any; Any -> Pro; Introduce hydrogen bonds; Hydrophobic packing; Cys-Cys bridges; Salt bridges; β-branched residues in β- strands; Pestering water from the core; etc. The main thing is that you should first know WHY the protein is unstable. Abstract: F U Applied: F LU I ©CMBI 2003
MUTATIONS ‘SHOULD’ ADD UP ©CMBI 2003
BUT THEY DON’T…. ©CMBI 2003
LOCAL UNFOLDING ©CMBI 2003
WEAK SPOTS IN PROTEINS ©CMBI 2003
WEAK SPOT PROTECTION ©CMBI 2003
SUPPORT FOR EXPERIMENTS • Selenomethionine for Xray; • Solubility (i.e. for NMR); • Tags for purification (His-tag, Flag-tag, etc); • Addition or removal of post-translational modification sites; • ‘Mutate away’ metal binding sites; • Introduce fluorophore; • Block binding, or make binding irreversible; • Etcetera. ©CMBI 2003
PREDICT MUTATIONS FROM ALIGNMENTS It is rapidly becoming apparent that multiple sequence alignments are the most powerful tool in bioinformatics. And that is also true for mutation design. If you can predict something that nature has done already, success is almost guaranteed. ©CMBI 2003
CONSERVED, VARIABLE, OR IN-BETWEEN QWERTYASDFGRGH QWERTYASDTHRPM QWERTNMKDFGRKC QWERTNMKDTHRVW Gray = conserved Black = variable Green = correlated mutations ©CMBI 2003
CORRELATED MUTATIONS SHAPE TREE 1 AGASDFDFGHKM 2 AGASDFDFRRRL 3 AGLPDFMNGHSI 4 AGLPDFMNRRRV ©CMBI 2003
CORRELATION = INFORMATION 1, 2 and 5 bind calcium; 3 and 4 don’t. Which residues bind calcium? 1 ASDFNTDEKLRTTYI Ca+ 2 ASDFSTDEKLKTTYI Ca+ 3 LSFFTTDTKLATIYI 4 LSHFLTDLKLATIYI 5 ASDFTTDEKLALTYI Ca+ ©CMBI 2003
AND NOW, THE VARIABLE RESIDUES 11 Red Main site 12 Orange Support 22 Yellow Communication 23 Green Modulator site 33 Blue The rest 20 Entropy at i: Ei = S pi ln(pi) i=1 Sequence variability is the number of residues that is present in more than 0.5% of all sequences. Entropy = Information Variability = Chaos Orange -> purple On this PC/beamer ©CMBI 2003
Entropy - variability 11 Red Main site 12 Orange Support 22 Yellow Communication 23 Green Modulator site 33 Blue The rest 20 Ei = S pi ln(pi) i=1 Entropy = Information Variability = Chaos Sequence variability is the number of residues that is present in more than 0.5% of all sequences. ©CMBI 2003
Entropy - Variability – Function* *This is for nuclear hormone receptors ©CMBI 2003
Acknowledgements V.G.H.Eijsink, B.v.d.Burg, G.Venema, B.Stulp, J.R.v.d.Zee, H.J.C.Berendsen, B.Hazes, B.W.Dijkstra, O.R.Veltman, B.v.d.Vinne, F.Hardy, F.Frigerio, W.Aukema, J.Mansfeld, R.Ulbrich-Hofmann, A.d.Kreij. ©CMBI 2003
A short break for a word from our sponsors Laerte Oliveira Adje Margot F L O R E N C E H O R N Our industrial sponsor: Wilma Kuipers Weesp Bob Bywater Copenhagen Nora vd Wenden The Hague Mike Singer New Haven Ad IJzerman Leiden Margot Beukers Leiden Fabien Campagne New York Øyvind Edvardsen TromsØ Simon Folkertsma Frisia Henk-Jan Joosten Wageningen Joost van Durma Brussels David Lutje Hulsik Utrecht Tim Hulsen Goffert Manu Bettler Lyon David Tim Elmar Krieger Fabien Manu Simon Folkertsma ©CMBI 2003