Download
1 / 1
10 likes | 99 Views
2. 5. Cotranslational Post-translational. Regulated Constitutive secretion secretion. Secretory vesicle. envelope. 3. Trans-Golgi network.

E N D
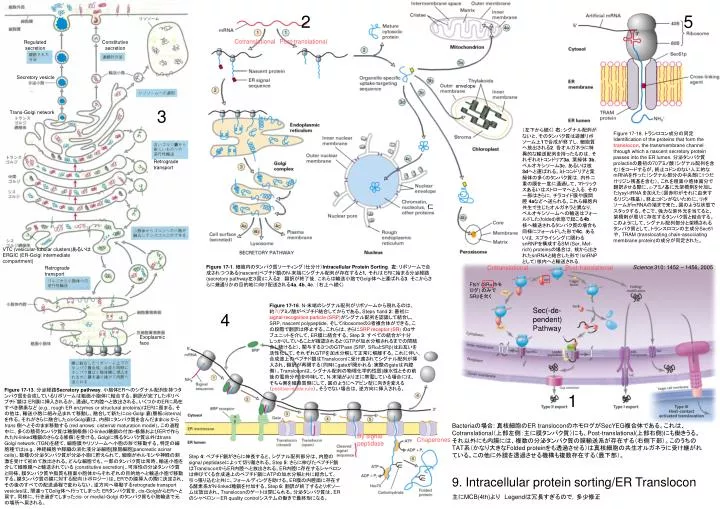
2 5 CotranslationalPost-translational Regulated Constitutive secretion secretion Secretory vesicle envelope 3 Trans-Golgi network (左下から続く) 右: シグナル配列がないと、そのタンパク質は遊離リボソーム上1で合成が終了し,細胞質へ放出される2.各オルガネラに特異的な輸送配列を持ったものは,それぞれミトコンドリア3a,葉緑体 3b,ペルオキシソーム3c,あるいは核 3dへと運ばれる。ミトコンドリアと葉緑体の多くのタンパク質は,内外二重の膜を一度に通過して、マトリックスあるいはストローマへと入る.その一部はさらに,チラコイド膜や膜間腔 4aなどへ送られる。これら細胞内共生で生じたオルガネラと異なり,ペルオキシソームへの輸送はフォールドした(folded)状態で起こる4b. 核へ輸送されるタンパク質の場合も同様にフォールドした形で4c,あるいは,スプライシングに関わるsnRNPを構成するSM (Ser, Met-rich) proteinsの場合は,核から出されたsnRNAと結合した形で(snRNPとして)核内へと輸送される. Figure 17-18. トランロコン成分の同定Identification of the proteins that form thetranslocon, the transmembrane channel through which a nascent secretory protein passes into the ER lumen. 分泌タンパク質prolactinの最初の70アミノ酸(シグナル配列を含む)をコードするが、終止コドンのない人工的なmRNAを作った(シグナル部分の中央部に1つだけリジン残基を含む)。これを粗面小胞体画分で翻訳させる際に、e-アミノ基に光架橋剤を付加したlysyl-tRNA を加えた(図赤印がそれに由来するリジン残基)。終止コドンがないために、リボソームがmRNAの端まで来た、図のような状態でスタックする。そこで、強力な紫外光を当てると、架橋剤が周りに存在するタンパク質と結合する。このようにして、シグナル配列部分と架橋されるタンパク質として、トランスロコンの主成分Sec61や、TRAM (translocating chain-associating membrane protein)の成分が同定された。 Retrograde transport 仁 VTC (vesicular-tubular clusters)あるいはERGIC (ER-Golgi intermediate compartment) Figure 17-1. 細胞内のタンパク質ソーティング(仕分け)Intracellular Protein Sorting.左: リボソームで合成されつつある(nascent)ペプチド鎖のN-末端にシグナル配列が存在すると1,それは ERに始まる分泌経路(secretory pathway左3図)に入る2 .翻訳が終了後,これらは輸送小胞でGolgi体へと運ばれる3.そこからさらに幾通りかの目的地に向け配送される4a, 4b, 4c.(右上へ続く) CotranslationalPost-translational Science 310: 1452 – 1456, 2005 Retrograde transport FtsY (SRaホモログ) のみでSRbを欠く Figure 17-16. N-末端のシグナル配列がリボソームから現れるのは、約70アミノ酸がペプチド結合してからである。Steps 1and 2: 最初に signal-recognition particle (SRP)がシグナル配列を認識して結合し、SRP, nascent polypeptide, そしてribosomeの3者複合体ができる。この段階で翻訳は停止する。これらは、さらにSRP receptor (SR)のαサブユニットを介して、ER膜に結合する。Step 3: すべての結合が十分しっかりしていることが確認されると(GTPが加水分解されるまでの間結合し続けると)、関与する3つのGTPase (SRP, SRaとSRb)はお互いを活性化して、それぞれGTPを加水分解して正常に解離する。これに伴い、合成途上のペプチド鎖はTransloconに受け渡されてシグナル配列が挿入され、翻訳が再開する(同時にgateが開かれる:実際のgateは内腔側)。Transloconは、シグナル配列の物理化学的性質(疎水性とその前後の電荷分布)を吟味して、N-末端がより正に帯電している場合には、そちら側を細胞質側にして、図のようにヘアピン型に向きを変える(positive-inside rule)。そうでない場合は、逆方向に挿入される。 Sec(-de-pendent) Pathway 4 Exoplasmic face Figure 17-13. 分泌経路Secretory pathway. 小胞体ERへのシグナル配列を持つタンパク質を合成しているリボソームは粗面小胞体に結合する。翻訳が完了したポリペプチド鎖は ER膜に挿入されるか、透過して内腔へと放出される。いくつかのERに局在すべき酵素など (e.g., rough ER enzymes or structural proteins)はERに留まる。その他は、輸送小胞に組み込まれて移動し、融合して新たにcis-Golgi 嚢(層板cisterna)を作る。それがさらに融合したcis-Golgi嚢は、内部にタンパク質を含んだままcisから trans側へとそのまま移動する (red arrows: cisternal maturation model)。この過程中に、多くの積荷タンパク質は糖鎖修飾(O-linked糖鎖の付加・修飾およびERで作られたN-linked糖鎖のさらなる修飾)を受ける。Golgiに残るタンパク質以外はtrans Golgi network (TGN)を経て、細胞膜やリソソームへ小胞の形で移動する。特定の細胞種では(e.g., 神経細胞や膵臓の消化液分泌細胞[腺房細胞]pancreatic acinar cells)、数種の分泌タンパク質が分泌小胞に貯えられて、細胞がホルモンや神経の刺激を受けて初めて放出される。どんな細胞でも、一部のタンパク質は常時、輸送小胞を介して細胞膜へと輸送されている (constitutive secretion)。可溶性の分泌タンパク質と同様、膜タンパク質や脂質も粗面小胞体からそれぞれの目的地へと輸送小胞で移動する。膜タンパク質の膜に対する配向(トポロジー)は、ERでの膜挿入の際に決定され、その後のすべての配送過程で変わらない。逆方向へ移動するretrograde transport vesiclesは、間違ってGolig体へ行ってしまった ERタンパク質を、cis-GolgiからERへと戻す。同様に、行き過ぎてしまったcis- or medial-Golgi のタンパク質も小胞輸送で元の場所へ戻される。 1 Bacteriaの場合: 真核細胞のER transloconのホモログがSecYEG複合体である。これは、Cotranslational(上部左側:主に膜タンパク質)にも、Post-translational(上部右側)にも働きうる。それ以外にも内膜には、複数の分泌タンパク質の膜輸送系が存在する(右側下部)。このうちのTAT系(かなり大きなFolded proteinをも透過させる)は真核細胞の共生オルガネラに受け継がれている。この他に外膜を透過させる機構も複数存在する(最下部)。 By signal peptidase Chaperones Step 4: ペプチド鎖がさらに伸長すると、シグナル配列部分は、内腔のsignal peptidaseによって切り離される。Step 5: さらに伸びたペプチド鎖はTransloconからER内腔へと放出される。ER内腔に存在するシャペロンは伸びてくる合成途上のペプチド鎖にATPの加水分解と共に結合して、引っ張り込むと共に、フォールディングを助ける。ER膜の内腔面に存在する酵素系がN-linked糖鎖を付加する。Step 6: 翻訳が終了するとリボソームは放出され、Transloconのゲートは閉じられる。分泌タンパク質は、ERのシャペロン-ER quality controlシステムの働きで最終形になる。 9. Intracellular protein sorting/ER Translocon 主にMCB(4th)より Legendは冗長すぎるので,多少修正






















