Download

1 / 19
200 likes | 291 Views
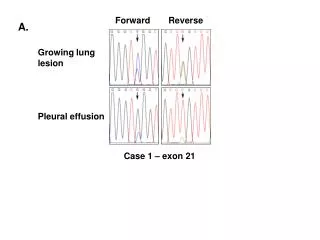
Forward Genetics Begins with a mutant (altered) phenotype and addresses the question, “what is the genotype?” or mutation in which gene caused the alteration in the phenotype. Reverse Genetics

E N D
Forward Genetics Begins with a mutant (altered) phenotype and addresses the question, “what is the genotype?” or mutation in which gene caused the alteration in the phenotype. Reverse Genetics Begins with the mutant gene sequence and asks the questions “what is the resulting effect on the phenotype?”
Genome sequencing has provided means to carry out systematic reverse genetics in large scale. Gene knock-out or null mutation provide direct means of studying the function of a gene product in situ. Arabidopsis is 120 Mb, contains small introns and small intergenic regions, therefore there is a high chance of finding mutation in every gene in T-DNA insertion population.
SATURATING THE GENOME WITH MUTATIONS Three variables determine the probability that a T-DNA insert will be found within a given gene: the size of the gene, the size of the genome, and the number of T-DNA inserts distributed among the population Curves are drawn for several different gene lengths: 0.5, 1, 2, 3, 4, and 5 kb. The probabilities were calculated using the following formula: P = 1 - (1 - [x/120,000])n, where P = probability of finding one T-DNA insert within a given gene, x = length of the gene in kilobases, and n = number of T-DNA inserts present in the population. This calculation assumes that the haploid Arabidopsis genome is 120 Mb and that T-DNA insertion is random.
Organization and Screening of 60,480 T-DNA–Transformed Arabidopsis Lines. (A) Pooling strategy. (B) Insertion screening strategy. 5' and 3' refer to PCR primers specific for the gene of interest. T-DNA L and T-DNA R refer to PCR primers specific for the T-DNA border regions. kanr, kanamycin resistant.
Alternatives to insertion mutations Why? First, many genes are functionallyredundant, sharing overlapping functions with other genes thatmay or may not be related at the sequence level. Mutation ofa functionally redundant gene is not likely to lead to an easilyrecognizable phenotype, because one or more other family memberscan provide the same function. Analysis of systematic gene knockoutshas revealed that a significant percentage of yeast genes haveno obvious phenotype when disrupted, despite testing under awide range of growth conditions. Therefore, it is likely that disruption of many plant geneswill not result in an easily identifiable phenotype. Second,many genes function at multiple stages of development. Mutationsin these genes may lead to early lethality or may be highlypleiotropic, which can mask the role of a gene in a specificpathway.
GUS Reporter Gene Expression Patterns in Representative Enhancer or Gene Trap Lines. Expression is evident, as follows: (A) In cotyledons and shoot apex but not leaves. (B) In trichomes. (C) In stipules and leaf tips. (D) In a single cell at the tip of leaf primordium. (E) In a lateral root primoridium. (F) In a root tip. (G) In a root cap.
Gene traps allow the identification of genesthat are not amenable to classical genetic analysis. Therefore,novel genes are likely to be found in any gene trap screen.Screens have in fact been successful in identifying genes specificallyexpressed in lateral roots, developingembryos,and shoot apices. Conditional screens havealso been performed to identify genes regulated by nematodeinfections andin response to abiotic stress. The PROLIFERA (PRL) gene was identified as a gene trap DsG insertionthat showed GUS activity in dividing cells. PRL encodes a proteinthat is related to MCM7, a member ofthe MCM gene family found in all eukaryotes and required forthe initiation of DNA replication. Expression in dividing cells is consistent withthis predicted role in cell division. Disruption of PRL by theDsG element led to megagametophyte and embryo lethality. Arrestof both megagametophytes and embryos occurred at variable stagesof development. There are many embryo-lethal mutations in Arabidopsis thathave variable phenotypes, which makes it difficult to determinethe cause of lethality. However, the GUS expressionpattern in dividing cells suggested a role in cell divisionbefore the gene was cloned.
Establishing an efficient Ac/Ds tagging system in rice: large-scale analysis of Ds flanking sequences Ac and Ds T-DNA vectors. RB and LB, right and left borders of T-DNA; 35S, CaMV 35S promoter; Ac, immobilized transposon; Ubi, maize ubiquitin promoter; gfp, green fluorescent protein; HPT, hygromycin phosphotransferase; gusA, -glucuronidase gene; bar, phosphinothricin acetyltransferase gene; Act1, rice actin promoter; I, intron from Arabidopsis G protein gene; A, triple splice acceptor; nos3' and ocs3', nopaline and octopine 3' UTR (poly(A) signal).
Large-scale generation of stable unlinked Ds transposants. Ds element was mobilized by crossing homozygous Ds and Ac lines; F1 seeds were planted and allowed to self-pollinate. In F1 generation, Ds transposed to a new locus in the genome, germinated F2 seeds were screened against GFP-linked donor sites, and the GFPseeds were planted on soil. At the three-leaf stage, plants were sprayed with Basta to select transposants. DNA was isolated from F2 plants, and bar-PCR was carried out to confirm the presence of Ds element.
Distribution of genetically mapped Ds sites The results indicated that different Ds elements showed preference for certain chromosomes: Ds3 (from chromosome 2) had a statistically significant preference for chromosome 7; Ds4 (from chromosome 1) for chromosome 6; Ds5 (from chromosome 8); and Ds9 (from chromosome 10) for chromosome 1
PTGS for functional genomics RNAi Injection of dsRNA into nematodes can trigger specific (homologous) RNA degradation. RNAi has been utilized to study 4000 genes in C. elegans. In plants, dsRNA and sshairpin RNA have similar effect. The predicted RNA structure and efficacy of gene-silencing constructs. (Ref: Wesley et al., 2001, Plant J. 27: 581-590) Constructs, controlled by Ubi1 promoter, silencing GUS in rice. Thick lines indicate a 560 nt GUS sequence; grey lines indicate non-GUS sequences; dashed grey lines indicate intron-junction sequences left after splicing; and short lines within the stem of hairpin structures indicate base pairing. Numbers in PTGS column indicate the percentage of plants showing GUS silencing; n = number of plants in each treatment.
From Ref: Wesley et al. (Ref: Wesley et al., 2001, Plant J. 27: 581-590) Using hpRNA constructs, we have obtained silenced plants for every gene that we targeted, irrespective of whether it was a viral gene, transgene or endogenous gene, and the silencing appears to be uniform within tissues in which the hpRNA is expressed. With ihpRNA constructs the efficiency averaged about 90%, and arms of 400±800 nt appear to be stable and effective. High levels of silencing were obtained with constructs having unmatched arm lengths, with arms as long as 853 nt or as little as 98 nt, and with arm sequences derived from coding, 3’ or 5’ untranslated regions of the target gene. These results suggest that ihpRNA constructs will be effective in a wide range of circumstances, and augur well for the generic use of the technology. The silencing was much more profound with ihpRNA constructs than either anti-sense or co-suppression constructs; some ihpRNA transformants were close to exhibiting a complete knockout of the target endogenous gene. However, most of the ihpRNA plants showed dramatically reduced but detectable levels of target gene activity.
VECTORS FOR VIRUS-INDUCED GENE SILENCING The regions of the plant that can be affected by gene silencing depend mostly upon viral vector characteristics. Silencing vectors usually travel systemically through the phloem in the vascular tissues to most parts of the plant. Although most of the viruses do not penetrate meristems, some have been found to deliver a silencing signal to meristematic regions of the plant. Some important characteristics of a viral silencing vector are its effectiveness in inducing silencing; its capability to infect and induce silencing in growing parts of the plant; its genome size (due to cloning steps for fragment insertion, since usually the smaller the vector the easier the cloning); its genome partition; the type of nucleic acid its genome is composed of (given that RNA plant viruses are the most common, though DNA viruses can facilitate inoculation as they allow the use of DNA instead of unstable RNA); its host range; whether its genome is or can be made available in binary vectors (to facilitate inoculation via Agrobacterium), and its safety for individuals and the environment
Tobacco mosaic virus (TMV) was the first viral vector used to successfully elicit VIGS of an endogenous gene in a plant species. Potato virus X (PVX) followed as the next viral vector used to carry a gene into a plant cell and trigger silencing via VIGS. Although effective, its incapability of infecting meristems is a great disadvantage, since delivering silencing to meristematic tissues is a sine qua non for studying genetic functions involved in developmental pathways. The advent of the Tobacco rattle virus (TRV) vector for VIGS opened the possibility of studying genes expressed in early organ development, because of its ability to reach growing points and to deliver the silencing signal to meristems. The cloning of its genome into binary Agrobacterium tumefaciens plasmids immensely facilitated the infection process
TRV is an ssRNA virus with a bipartite genome. The component called RNA1 encodes, among other genes, an RdRP, whereas its genome partner, called RNA2, encodes the coat protein. The target gene fragment for silencing is inserted into the RNA2 element. Inoculation, either mechanical or via agroinfiltration, requires the presence of both genome components. In the case of agroinfiltration, two different Agrobacterium clones, one carrying the RNA1 genome and another with the RNA2 containing the target gene fragment, are mixed together and co-infiltrated into the leaf tissues.
A significant limitation of the VIGS system is that TRV, and most of the other vectors available, work very well in N. benthamiana and in the tomato, but they do not show efficacy in Arabidopsis, in which a myriad of genes still need to be functionally analyzed. Additionally, a vector with the capability to infect monocot species, such as maize, rice and lily, is also greatly desired.
Silencing of two endogenous genes was achieved from DNA fragments carried in different TGMV component vectors. Variegation occurred in leaves that were partly expanded at the time of inoculation, however, very little stem elongation was evident in new growth (k). Plant (l) inoculated with A::790su/B::122PCNA and pruned (arrow) 2 weeks after inoculation showed silencing in axillary buds. PCNA silencing is evidenced by reduced stem elongation and aberrant leaf formation. The two axillary outgrowths show different degrees of su silencing with one cluster of leaves (right) showing almost no chlorophyll. Note circular yellow spots in inoculated lower leaves (black arrow). Silencing of a meristematic gene using geminivirus-derived vectors
Silencing of the PDS gene. Infection of recombinant TRV carrying the PDS (phytoene desaturase) sequence silences endogenous PDS in N. benthamiana plants and causes inhibition of carotenoid biosynthesis resulting in the photobleaching phenotype. On the left, whole plant and on the right, enlarged single leaf.