Download

1 / 34
340 likes | 945 Views
Clonaggio posizionale. Con questo termine si indica l’insieme delle metodiche che possono essere impiegate per identificare un gene utilizzando come informazione di partenza solo la sua posizione fisica sul cromosoma. Le informazioni sulla posizione del gene possono derivare da:

E N D
Clonaggio posizionale • Con questo termine si indica l’insieme delle metodiche che possono essere impiegate per identificare un gene utilizzando come informazione di partenza solo la sua posizione fisica sul cromosoma. • Le informazioni sulla posizione del gene possono derivare da: • Analisi di linkage • Anomalie cromosomiche in alcuni pazienti (delezioni, traslocazioni) • Perdita di eterozigosi per marcatori polimorfici (importante soprattutto per l’identificazione dei geni oncosoppressori)
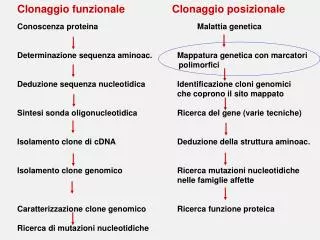
Fasi del clonaggio posizionale • Prima identificazione della regione cromosomica (di solito identifica regioni di circa 10 megabasi) • Restringimento della regione candidata (per poter procedere bisogna che il gene sia mappato in una regione di circa una megabase) • Mappaggio e sequenziamento della regione candidata • Identificazione dei geni situati nella regione critica, e ricerca di quelli che presentano anomalie. • Conferma del gene • In seguito al sequenziamento dl genoma umano questa procedura si è semplificata moltissimo, e oggi, dopo aver mappato il gene, si può in genere procedere direttamente alla sua identificazione.
Anomalie citogenetiche Traslocazioni Delezioni
Perdita di eterozigosi A’ B’ C’ A’’ B’’ C’’ A’ B’ C’ A’’ C’’ Allele mutato Allele normale Cellula affetta Cellula normale A seguito della delezione la cellula è diventata apparentemente omozigote al locus B
Perdita di eterozigosi La cellula diventa anormale perché un allele era già mutato, mentre il secondo viene perso a causa della delezione. A seguito di questa delezione la cellula è diventata apparentemente omozigote al locus B. Il locus B non è responsabile della patologia. Tuttavia, se rilevo questo fenomeno (confrontando il tessuto sano e quello patologico dello stesso individuo per i diversi marcatori della regione), capisco che il gene si trova vicino al locus B.
Identificazione del gene della distrofia muscolare di Duchenne
Identificazione del gene della distrofia muscolare di Duchenne • Malattia a trasmissione X-linked recessiva • In genere colpiti i maschi, ma raramente anche le femmine • 1/3 dei casi sono nuove mutazioni • Gene mappato su Xp21 mediante analisi di linkage già nel 1982 (primo gene ad essere mappato con questo metodo). • Prima della identificazione del gene non si aveva alcuna idea sulle basi biochimiche della patologia. • Il lavoro che ha portato alla identificazione di questo gene (1987) ha costituito il primo banco di prova del clonaggio posizionale. Due gruppi sono arrivati indipendentemente alla identificazione del gene, in entrambi i casi usando scorciatoie rispetto all’approccio classico.
Strategia 1: Clonaggio per sottrazione In questo caso si è sfruttato un paziente che presentava una delezione su Xp21 visibile citogeneticamente. Soggetto normale Paziente
Strategia 1: Clonaggio per sottrazione DNA soggetto normale: Taglio con enzima A DNA paziente: Taglio con enzima B
Strategia 1: Clonaggio per sottrazione Il DNA del soggetto normale vine mescolato con un eccesso di DNA del paziente e denaturato. La miscela viene poi fatta rinaturare.
Strategia 1: Clonaggio per sottrazione Situazione dopo rinaturazione
Strategia 1: Clonaggio per sottrazione Solo le molecole di DNA provenienti dalla regione critica non ibridizzano con le molecole di DNA del paziente e quindi mantengono le estremità coesive caratteristiche dell’enzima A A A A A A A Pertanto queste molecole possono essere inserite selettivamente in vettori di clonaggio tagliati con l’enzima A. I risultanti cloni provengono preferenzialmente dalla regione critica, e possono quindi essere utilizzati per avvicinarsi al gene
Strategia 2: Clonaggio del punto di rottura di una traslocazione in una femmina affetta Le femmine sono colpite dalla DM di Duchenne rarissimamente (circa 20 casi al mondo). Generalmente si tratta di casi sporadici (senza storia familiare) in cui sono presenti traslocazioni tra cromosoma X e autosomi.
Strategia 2: Clonaggio del punto di rottura di una traslocazione in una femmina affetta Nel caso che ha portato all’isolamento del gene la paziente presentava una traslocazione tra il cromosoma X (nella regione Xp21) e il cromosoma 21. La regione del cromosoma 21 coinvolta era nota contenere un gruppo di geni codificanti per RNA ribosomiali (circa 50 ripetizioni). Pertanto, per clonare il punto di rottura, la strategia è stata di produrre una genoteca con il DNA genomico della paziente (library genomica) e isolare i cloni che contenevano contemporaneamente geni per l’RNA ribosomiale, e materiale proveniente dal cromosoma X.
Entrambi gli approcci hanno portato alla identificazione dello stesso gene • Si tratta di un gene enorme: • 2,4 milioni di basi • 80 esoni • Moltissimi prodotti derivanti da promotori multipli e splicing alternativo. • La maggior parte dell mutazioni sono delezioni • Lo stesso gene è anche coinvolto nella distrofia muscolare di Becker (forma meno grave derivante da mutazioni meno devastanti).
Fibrosi cistica del pancreas • Patologia autosomica recessiva, molto frequente • Anomala viscosità delle secrezioni, che determina chiusura dei dotti pancreatici (maldigestione) e dei bronchi (polmoniti recidivanti e bronchiectasie). • Anomala concentrazione di cloro nelle secrezioni (test del sudore) Pancreas normale Pancreas di paziente
Identificazione del gene della fibrosi cistica • Nel 1985 il gene venne localizzato sul cromosoma 7 grazie al suo linkage con un marcatore proteico. • Successivamente fu possibile un mappaggio più fine, che localizzò il gene in un intervallo di 500 kb tra due marcatori (cMet e D7S8). • Il DNA di tutta la regione è stato clonato usando le tecniche del chromosome walking e jumping. • Il sequenziamento di questi cloni ha permesso di identificare nuovi marcatori polimorfici, in forte linkage disequilibrium con il gene della fibrosi cistica. Questo è stato possibile perché la maggior parte degli alleli mutati nella popolazione non derivano da nuove mutazioni, ma da un’unica mutazione ancestrale, mantenuta in virtù del vantaggio dell’eterozigote (gli eterozigoti sono probabilmente più resistenti del resto della popolazione all’infezione da Salmonella typhi).
Sonda 1 Sonda 2 Sonda 3 Sonda 4 Come si fa a clonare una regione genomica partendo da una sonda corrispondente ad un punto per il quale sia dsponibile una sonda molecolare? Gene A E C F D B G Questa metodica prende il nome di chromosome walking
Camminare e saltare lungo il cromosoma con diversi tipi di cloni Marcatore 1 Regione critica 2 Mb Marcatore 2 Gene malattia Cloni YAC (1Mb) Cloni BAC, P1, PAC (100-200 Kb) Cloni fagici e cosmidici (20-40 Kb)
Linkage disequilibrium Alleli marcatori Cromosomi FC Cromosomi normali X1, K1 3 49 X1, K2 147 19 X2, K1 8 70 X2, K2 8 25 A1 X1 K2 B3 A X FC K B A2 X2 K1 B1
Identificazione del gene della fibrosi cistica • Il sequenziamento della regione fiancheggiante i marcatori più vicini al gene della fibrosi cistica permise di identificare il primo esone del gene. • Il resto è stato identificato facendo diversi screening di cDNA libraries. • Alla fine nei pazienti è stata trovata nel 70% dei casi una mutazione in omozigosi consistente nella delezione di un codone (F508Del). • Non è stato facile stabilire che questa mutazione è effettivamente causa della malattia, poiché si trova nel 3-4% della popolazione normale. Avrebbe potuto essere un ulteriore polimorfismo strettamente associato al gene. Tuttavia in seguito sono state riscontrate altre mutazioni nello stesso gene in altri pazienti.
Identificazione del gene della fibrosi cistica • Oltre a ciò si è visto che la proteina codificata da questo gene è un canale di membrana per gli ioni cloro, e che la mutazione disturba la sua capacità di trasportare ioni.
Identificazione dei geni oncosoppressori mediante approcci posizionali • I soggetti predisposti ad alcune forme di cancro sono spesso portatori di mutazioni in eterozigosi a carico di geni oncosoppressori • Una mutazione somatica (ad esempio una delezione) che determini una inattivazione della seconda copia del gene in una o più cellule può determinare l’insorgenza del cancro. • Se la delezione è grande, la si vede anche al microscopio, e questo può servire per dare le prim indicazioni sulla posizione del gene. • Se la delezione è piccola, nel tessuto tumorale del paziente si nota una perdita di eterozigosi per i marcatori associati al gene. • L’analisi di molti tumori permette di restringere • la regione critica e di mappare finemente la • posizione del gene oncosoppressore
A’ B’ C’ A’’ B’’ C’ A’’ B’’ C’’ A’’’ B’ C’’’ A’ B’ C’ A’’ B’’ C’ A’’ C’’ A’’’ B’ Paziente 1 Paziente 2 Tumore Tumore Sangue Sangue Genotipo Genotipo A’ A’’ B’ B’’ C’ C’’ A’ A’’ B’ C’ C’’ A’’ A’’’ B’ B’’ C’ C’’’ A’’ A’’’ B’ B’’ C’ Conclusione: il gene è localizzato tra il locus B e il locus C
In seguito al sequenziamento del genoma umano non è più necessario fare tutto il lavoro di mappaggio e sequenziamento delle regioni critiche. Se identifico la regione critica di un gene malattia la trovo già mappata e sequenziata, e so anche quali geni sono compresi in essa.
Approccio del gene candidato indipendente dalla posizione genomica Si tratta di una situazione abbastanza rara. Il caso più tipico è quello della identificazione di un gene malattia umano basata sulle omologie con un modello sperimentale animale. Se nel modello viene identificato un gene responsabile di una patologia che ricorda molto una patologia umana, il gene omologo umano diventa immediatamente il candidato più forte per quella patologia. Ovviamente il candidato andrà confermato cercando mutazioni di quel gene nei pazienti.
Approccio del candidato posizionale • La maggior parte dei geni responsabili di patologie umane sono stati identificati grazie ad una combinazione di informazioni posizionali e non posizionali. • Infatti bisogna considerare che: • Un approccio completamente indipendente dalla posizione funziona solo raramente, perché molto spesso le basi molecolari delle malattie sono così complicate che non è possibile fare ipotesi funzionali sensate. Infatti molto spesso, dopo l’identificazione di un gene-malattia, si è scoperto che le ipotesi sulla sua funzione erano completamente sbagliate. • Un approccio puramente posizionale non è quasi mai efficace, perchè le regioni candidate in genere contengono decine di geni, e cercare sistematicamente le mutazioni in tutti è un lavoro troppo grosso. • Pertanto, anche quando si usa il clonaggio posizionale, e importante utilizzare una serie di altri criteri per identificare i candidati migliori.
Criteri per la definizione di un candidato posizionale • Espressione compatibile con la patologia • Funzione appropriata (esempio della rodopsina) • Conservazione evolutiva (DRES) • Omologia con un gene implicato in un modello animale della malattia • (sindrome di Waardenburg e topo Splotch)
Il sequenziamento del genoma umano e le nuove tecnologie rendono estremamente più semplice l’approccio del candidato posizionale Sindrome di Leigh, tipo Franco-Canadese • Patologia piuttosto rara, ma frequente in una particolare area geografica. • Degenerazione di alcune aree cerebrali (nuclei della base e tronco dell’encefalo), associata a episodi ripetuti di acidosi lattica. • Deficit di Citocromo c ossidasi • La citocromo c ossidasi è un complesso mitocondriale costituito da almeno 30 subunità; il suo assemblaggio è regolato da altre proteine
L’analisi di linkage aveva permesso di restringere la regione critica in un intervallo di 5 cM sul cromosoma 2. • La mappa del genoma ha permesso immediatamente di identificare quali geni sono presenti in questa regione (15 verificati sperimentalmente e 15 predetti da approcci informatici • Nessun gene mitocondriale ovvio tra questi • Una analisi di dati di microarray ha permesso di stabilire che uno di questi geni (LRPPRC) ha un profilo di espressione simile a quello dei geni mitocondriali noti. Infatti l’analisi dei pazienti ha evidenziato mutazioni proprio a carico di questo gene.
Conferma del gene candidato • Identificazione di mutazioni nei pazienti (si veda la lezione sulla patologia molecolare) e analisi della loro segragazione nelle famiglie affette • Produzione di un modello sperimentale animale della malattia mediante inattivazione selettiva (knockout) o overespressione del gene omologo in animali transgenici. • Se possibile, la prova migliore del coinvolgimento diretto di un gene in una patologia consiste nella restaurazione del fenotipo normale in un modello cellulare o animale della stessa (saggio di complementazione). • Una volta identificato il gene bisogna capire quali sono i meccanismi fisiologici in cui è coinvolto e quali sono gli effetti funzionali delle mutazioni.