Download

1 / 33
350 likes | 1.24k Views
Biochemistry 412 Enzyme Kinetics II April 1 st , 2005. Acknowledgement: For this lecture, too, I am greatly indebted to Dr. Gabriel Fenteany, Department of Chemistry, University of Illinois at Chicago, for many of the slides. k cat , K M , and k cat /K m : Catalytic Efficiency.

E N D
Biochemistry 412 Enzyme Kinetics II April 1st, 2005
Acknowledgement: For this lecture, too, I am greatly indebted to Dr. Gabriel Fenteany, Department of Chemistry, University of Illinois at Chicago, for many of the slides.
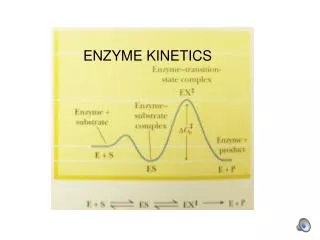
kcat, KM, and kcat/Km: Catalytic Efficiency Substrate preferences for chymotrypsin Note
For [S] < Kmv ≈ Vmax[S]/Km = kcat[E]t[S]/Km= [E]t(kcat/Km)[S]
Why is the kcat/Km ratio a key measure of catalytic efficiency?The answer lies with the free energy of the enzyme:transition state complex. The free energy of this state has to be low enough to enable the enzyme to bind the substrate at physiological concentrations, but high enough not to unduly restrict k2(kcat).
Note: with certain industrial processes involving highly soluble substrates (e.g., fructose production by xylose isomerase) where the reaction can be run at substrate concentrations much above the Km of the natural enzyme, you can readily improve the productivity of the enzymatic reaction by mutating the enzyme to increase both Km and kcat in tandem (keeping kcat/Km approximately constant).
Protein engineering to change the cofactor specificity of 2,5-DKG reductase Banta et al (2002)
S1 + S2 P1 + P2 A-X + B A + B-X (in transferase reactions) Sequential binding of S1 and S2 before catalysis: Random substrate binding - Either S1 or S2 can bind first, then the other binds. Ordered substrate binding - S1 must bind before S2. Ping Pong reaction - first S1 P1, P1 released before S2 binds, then S2 P2. Bisubstrate Reactions E E
E + S ES EP2 E + P2 The Pre-Steady State in Chymotrypsin-Catalyzed Hydrolysis of p-Nitrophenyl Acetate* H2O P1 k1 k2 k3 k-1 v = kcat[E]t[S]/(KM + [S]) Steady-state velocity, where kcat = k2k3/(k2 + k3) KM =KSk3/(k2 + k3) KS = k-1/k1 For chymotrypsin: k2 >> k3 Release of P1 is faster than EP2 breaking down to E + P2 *Note: such substrates are called “burst titrants” because they turn over rapidly to release chromophore only once. By back-extrapolation, they can be used to quantitate the amount of active enzyme in a reaction solution.
Reversible inhibition (inhibitors that can reversibly bind and dissociate from enzyme, activity of enzyme recovers when inhibitor diluted out, usually non-covalent interaction) Competitive Mixed (noncompetitive) Uncompetitive Irreversible inhibition (inactivators that irreversibly associate with enzyme, activity of enzyme does not recover with dilution, usually covalent interaction) Types of Enzyme Inhibition
Lineweaver-Burk plots illustrating different modes of inhibition
Competitive Inhibition Typically, I is a substrate analog.
Effects of Competitive Inhibitor on Enzyme Kinetics KI (inhibitor dissociation constant)=koff/kon KappM = KM(1 + [I]/KI)> KM Vappmax = Vmax
Effects of Mixed (Noncompetitive) Inhibitor on Enzyme Kinetics These inhibitors affect kcat only. KappM = KM Vappmax = Vmax/(1 + [I]/KI) < Vmax
Effects of Uncompetitive Inhibitor on Enzyme Kinetics • Not the same as noncompetitive (mixed) inhibition. • In uncompetitive inhibition, inhibitor only binds ES and not E alone. KappM = KM/(1 + [I]/KI)< KM Vappmax = Vmax/(1 + [I]/KI)< Vmax
Irreversible Inhibition k1 k2 E + I E·I E-I Plot: ln(residual enzyme activity) vs. time If [I]>>[E], conditions are pseudo-first order and slope is -kobs (pseudo-first order inactivation rate constant) kinact (second-order inactivation constant) = k1k2/k-1 = kobs/[I] k-1 Slope = -kobs
Irreversible Inhibition by Adduct Formation (diisopropylfluorophosphate)
Irreversible Inhibition of Chymotrypsin by TPCK (N-tosyl-L-phenylalanine chloromethylketone)
Final Topic Q: What can you learn by “de-fanging” an enzyme by mutating out its catalytic residues?
Reaction mechanism of a serine protease (in this case, subtilisin) Note the three residues of the “catalytic triad”: Ser221, His64, & Asp32.
Take home lesson: even with no catalytic residues, the enzyme still accelerates the reaction better than 1000-fold the rate of the uncatalyzed reaction.Way to bind that transition state!