Download

1 / 19
280 likes | 1.98k Views
Biosynthesis of Fatty Acids. Medical Biochemistry Lecture #46. FattyAcids. Fatty acids are a class of compounds containing a long hydrocarbon chain and a terminal carboxylate group. Nomenculature:

E N D
Biosynthesis of Fatty Acids Medical Biochemistry Lecture #46
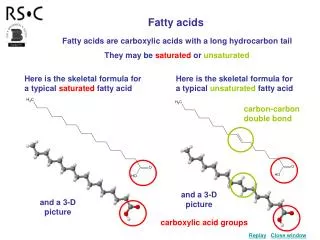
FattyAcids Fatty acids are a class of compounds containing a long hydrocarbon chain and a terminal carboxylate group. Nomenculature: - Systematic name for a fatty acid is derived from the name of its parent hydrocarbon by the substitution of oic for the final e. - For example, the C18 saturated fatty acid is called octadecanoic acid (18:0) because the parent hydrocarbon is octadecane.
F.A. Nomenclature (cont.) - C18 with one double bond is called octadecenoic acid (18:1); with two double bonds is called octadecadienoic acid (18:2); with three double bonds, octadecatrienoic acid (18:3).
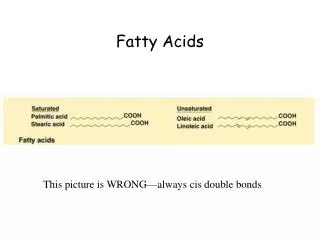
Fatty acids vary in chain length and degree of unsaturation: • Usually contain an even number of carbon atoms, typically between 14 and 24. The 16- and 18-carbon fatty acids are most common. • May contain one or more double bonds. The double bonds in polyunsaturated fatty acids are separated by at least one methylene group. • The configuration of the double bonds in most unsaturated fatty acids is cis.
Properties of fatty acids are markedly dependent on their chain length and on the degree of saturation. • -Melting point of stearic acid is 69.6oC, whereas that of oleic acid (with one double bond) is 13.4oC. • –Melting temperature of palmitic acid (C16) is 6.5 degrees lower than that of stearic acid (C16)
FATTY ACID SYNTHESIS (LIPOGENESIS) • Glucose provides the primary substrate for lipogenesis • In humans, adipose tissue may not be an important site, and liver has only low activity • Variations in fatty acid synthesis between individuals may have a bearing on the nature and extent of obesity, and one of the lesions in type I, insulin-dependent diabetes mellitus is inhibition of lipogenesis DE NOVO SYNTHESIS OCCURS IN CYTOSOL • Liver, kidney, brain, lung, mammary gland, and adipose tissue.
Step 1: Formation of Malonylcoenzyme A is the committed step in fatty acid synthesis: It takes place in two steps: carboxylation of biotin (involving ATP) and transfer of the carboxyl to acetyl-CoA to form malonyl-CoA. Reaction is catalyzed by acetyl-CoA carboxylase. It is a multienzyme protein. The enzyme contains a variable number of identical subunits, each containing biotin, biotin carboxylase, biotin carboxyl carrier protein, and transcarboxylase, as well as a regulatory allosteric site.
Step 2: • Fatty acid synthase catalyzes the remaining steps. It is a multienzyme polypeptide complex that contains acyl carrier protein (ACP). ACP contains the vitamin pantothenic acid in the form of 4'-phosphopantetheine. ACP takes over the role of CoA. • It offers great efficiency and freedom from interference by competing reactions • Synthesis of all enzymes in the complex is coordinated, since it is encoded by a single gene • It is a dimer, and each monomer is identical, consisting of one chain containing all seven enzyme activities of fatty acid synthase and an ACP with a 4'-phosphopantetheine-SH group. Dimer is arranged in a "head to tail" configuration. Monomer is not active.
Step 3: Elongation of fatty acid chains occurs in endoplasmic reticulum • This pathways "microsomal system" converts fatty acyl-CoA to an acyl-CoA derivative having two carbons more, using malonyl-CoA as acetyl donor and NADPH as reductant catalyzed by the microsomal fatty acid elongase system of enzymes.
Nutritional state regulates lipogenesis: • Lipogenesis converts surplus glucose and intermediates such as pyruvate, lactate, and acetyl-CoA to fat. • Rate is higher in well-fed animals whose diets contains a high proportions of carbohydrates. • It is depressed under conditions of restricted caloric intake, on a high-fate diet, or when there is a deficiency of insulin, as in diabetes mellitus. All these conditions are associated with increased concentrations of plasma free fatty acids. • There is an inverse relationship between hepatic lipogenesis and the concentration of serum-free fatty acids. The greatest inhibition of lipogenesis occurs over the range of free fatty acids (0.3-0.8 µmol/mL pf plasma).
Fat in the diet also causes depression of lipogenesis in the liver, and when there is more than 10% of fat in the diet, there is little conversion of dietary carbohydrates to fat.
SHORT AND LONG-TERM MECHANISMS REGULATE LIPOGENESIS • In the short-term, synthesis is controlled by allosteric and covalent modification of enzymes; For long-term, there are changes in gene expression Short-term • Acetyl-CoA carboxylase is most important in regulating synthesis • Activated by citrate, which increases in well-fed state and is an indicator of a plentiful supply of acetyl-CoA • Inhibited by long-chain acyl-CoA. • Pyruvate dehydrogenase regulates availability of free acetyl-CoA for lipogenesis. Acetyl-CoA causes an inhibition of pyruvate dehyrogenase.
Hormones (short term) • Insulin stimulates lipogenesis by several mechanisms: • a. increases transport of glucose into the cell (e.g., adipose tissues) and thereby increases the availability of both pyruvate for fatty acid synthesis and glycerol-3-phosphate for esterification of the newly formed fatty acids. • b. Converts inactive form of pyruvate dehydrogenase to the active form in adipose tissues • c. Activates acetyl-CoA carboxylase • d. Insulin depress intracellular cAMP levels, inhibits lipolysis • e. Insulin antagonizes the actions of glucagon and epinephrine
Long-term • Expression is increased in response to fed state and is decreased in fasting, feeding of fat, and in diabetes (adaptive mechanism).