Download
1 / 1
10 likes | 177 Views
FUNCTIONAL SPECIALIZATION OF HUMAN AUDITORY CORTICAL FIELDS G. C. Stecker 1,2 , I. Liao 1 , T.J.Herron 1 , X.Kang 1 , E.W.Yund 1 , T. J. Rinne 1,3 , D.L.Woods 1 *

E N D
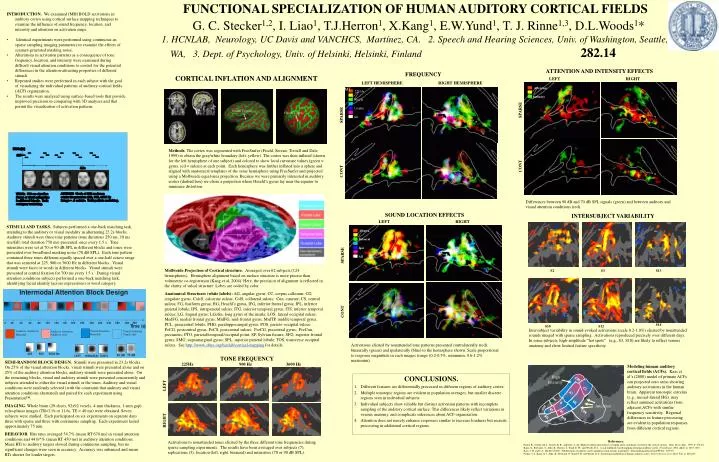
FUNCTIONAL SPECIALIZATION OF HUMAN AUDITORY CORTICAL FIELDS G. C. Stecker1,2, I. Liao1, T.J.Herron1, X.Kang1, E.W.Yund1, T. J. Rinne1,3, D.L.Woods1* 1. HCNLAB, Neurology, UC Davis and VANCHCS, Martínez, CA. 2. Speech and Hearing Sciences, Univ. of Washington, Seattle, WA, 3. Dept. of Psychology, Univ. of Helsinki, Helsinki, Finland 282.14 HG HG HG S2 S3 S13 HG HG HG FREQUENCY SOUND LOCATION EFFECTS LEFT LEFT RIGHT RIGHT Contra Auditory Cortex S14 S10 S12 Binaural Ipsi All AC AC Heschl’s gyrus SPARSE Heschl’s gyrus MT MT IPL IPL SPARSE LO LO V3a V3a CONT CONT LEFT HEMISPHERE RIGHT HEMISPHERE MT 225Hz Activations elicited by nonattended tone patterns presented contralaterally (red), binaurally (green) and ipsilaterally (blue) to the hemisphere shown. Scale proportional to response magnitude in each images (range (0.2-0.5%, minimum, 0.6-1.2% maximum). 900Hz 3.6 kHz Differences between 90 dB and 70 dB SPL signals (green) and between auditory and visual attention conditions (red). SPARSE Attention ATTENTION AND INTENSITY EFFECTS INTERSUBJECT VARIABILITY All Intensity CONT Intersubject variability in sound-evoked activations (scale 0.2-1.0%) elicited by nonattended sounds imaged with sparse sampling. Activations reproduced precisely over different days. In some subjects, high-amplitude “hot spots” (e.g., S3, S10) are likely to reflect venous anatomy and show limited feature specificity. Attention TONE FREQUENCY 225Hz 900 Hz 3600 Hz High Intensity LEFT RIGHT Activations to nonattended tones elicited by the three different tone frequencies during sparse sampling experiments. The results have been averaged over subjects (7), replications (3), location (left, right, binaural) and intensities (70 or 90 dB SPL). • INTRODUCTION. We examined fMRI BOLD activations in auditory cortex using cortical surface mapping techniques to examine the influence of sound frequency, location, and intensity and attention on activation maps. • Identical experiments were performed using continuous an sparse sampling imaging parameters to examine the effects of scanner-generated masking noise. • Alterations in activation patterns as a consequence of tone frequency, location, and intensity were examined during difficult visual attention conditions to control for the potential differences in the attention-attracting properties of different stimuli. • Repeated studies were performed in each subject with the goal of visualizing the individual patterns of auditory cortical fields (ACF) organization. • The results were analyzed using surface-based tools that provide improved precision in comparing with 3D analyses and that permit the visualization of activation patterns CORTICAL INFLATION AND ALIGNMENT AC Methods. The cortex was segmented with FreeSurfer (Fischl, Sereno, Tootell and Dale, 1999) to obtain the gray/white boundary (left, yellow). The cortex was then inflated (shown for the left hemisphere of one subject) and colored to show local curvature values (green = gyrus, red = sulcus) at each point. Each hemisphere was further inflated into a sphere and aligned with anatomical templates of the same hemisphere using FreeSurfer and projected using a Mollweide equal-area projection. Because we were primarily interested in auditory cortex (dashed box) we chose a projection where Heschl’s gyrus lay near the equator to minimize distortion. STIMULI AND TASKS. Subjects performed a one-back matching task, attending to the auditory or visual modality in alternating 23.2s blocks. Auditory stimuli were three-tone patterns (tone durations 250 ms, 10 ms rise/fall, total duration 750 ms) presented once every 1.5 s. Tone intensities were set at 70 or 90 dB SPL in different blocks and tones were presented over broadband masking noise (70 dB SPL). Each tone pattern contained three tones different equally spaced over a one-half octave range that was centered at 225, 900 or 3600 Hz in different blocks. Visual stimuli were faces or words in different blocks. Visual stimuli were presented at central fixation for 700 ms every 1.5 s. During visual attention conditions subjects performed a one-back matching task, identifying facial identity (across expressions) or word category. Mollweide Projection of Cortical structure. Averaged over 62 subjects (124 hemispheres). Hemisphere alignment based on surface structure is more precise than volumetric co-registration (Kang et al, 2004). Here, the precision of alignment is reflected in the clarity of sulcal structure. Lobes are coded by color. Anatomical Structures (white labels): AG, angular gyrus; CC, corpus callosum; CG, cingulate gyrus; CalcS, calcarine sulcus; ColS, collateral sulcus; Cun, cuneate; CS, central sulcus; FG, fusiform gyrus; HG, Heschl’s gyrus, IFG, inferior frontal gyrus; IPL, inferior parietal lobule; IPS, intraparietal sulcus; ITG, inferior temporal gyrus; ITS, inferior temporal sulcus; LG, lingual gyrus; LGofin, long gyrus of the insula; LOS, lateral occipital sulcus; MedFG, medial frontal gyrus; MidFG, mid-frontal gyrus, MidTP, middle temporal gyrus; PCL, paracentral lobule; PHG, parahippocampal gyrus; POS, parieto-occipital sulcus; PoCG, postcentral gyrus, PoCS, postcentral sulcus; PreCG, precentral gyrus; PreCun, precuneus; PTO, parietal/temporal/occipital point; SF, Sylvian fissure; SFG, superior frontal gyrus; SMG, supramarginal gyrus; SPL, superior parietal lobule; TOS, transverse occipital sulcus. See http://www.ebire.org/hcnlab/cortical-mapping for details. SEMI-RANDOM BLOCK DESIGN. Stimuli were presented in 23.2s blocks. On 25% of the visual attention blocks, visual stimuli were presented alone and on 25% of the auditory attention blocks, auditory stimuli were presented alone. On the remaining blocks, visual and auditory stimuli were presented concurrently and subjects attended to either the visual stimuli or the tones. Auditory and visual conditions were randomly selected (with the constraint that auditory and visual attention conditions alternated) and paired for each experiment using Presentation™. IMAGING. Whole brain (29-slices, 92x92 voxels, 4-mm thickness, 1-mm gap) echo-planar images (TR=2.9s or 11.6s, TE = 40 ms) were obtained. Seven subjects were studied. Each participated on six experiments on separate days three with sparse and three with continuous sampling. Each experiment lasted approximately 75 min. BEHAVIOR. Hits rates averaged 54.7% (mean RT 670 ms) in visual attention conditions and 44.0/^% (mean RT 450 ms) in auditory attention conditions. Mean RTs to auditory targets slowed during continuous sampling, but no significant changes were seen in accuracy. Accuracy was enhanced and mean RTs shorter for louder targets. Modeling human auditory cortical fields (ACFs). Kaas et al’s (2000) model of primate ACFs can projected onto areas showing auditory activations in the human brain. Apparent tonotopic extrema (e.g., mesial-lateral HG) may reflect summed activations from adjacent ACFs with similar frequency sensitivity. Regional differences in feature processing are evident in population responses from different cortical regions. • CONCLUSIONS. • Different features are differentially processed in different regions of auditory cortex. • Multiple tonotopic regions are evident in population averages, but smaller discrete regions seen in individual subjects. • Individual subjects show reliable but distinct activation patterns with incomplete sampling of the auditory cortical surface. This differences likely reflect variations in venous anatomy, and complicate inferences about ACF organization. • Attention does not merely enhance responses similar to increase loudness butrecruits processing in additional cortical regions. References. Fischl, B., Sereno, M. I., Tootell, R. B., and Dale, A. M. High-resolution intersubject averaging and a coordinate system for the cortical surface. Hum. Brain Map., 1999, 8: 272-84. Kang, X., Bertrand., 0., Alho, K., Herron, T., Yund, E. W., and Woods, D. L. Local landmark-based mapping of human auditory cortex.NeuroImage, 2004. 22(4): p. 1657-1670. Kaas, J. H. and T. A. Hackett (2000). "Subdivisions of auditory cortex and processing streams in primates." Proc Natl Acad Sci U S A97(22): 11793-9. Petkov, C.I., Kang, X.J., Alho, K., Bertrand, O. Yund, E.W. and Woods, D.L. Attentional modulation of human auditory cortex. Nature Neuroscience 2004. 7(6): p. 658-663.