Download
1 / 1
10 likes | 186 Views
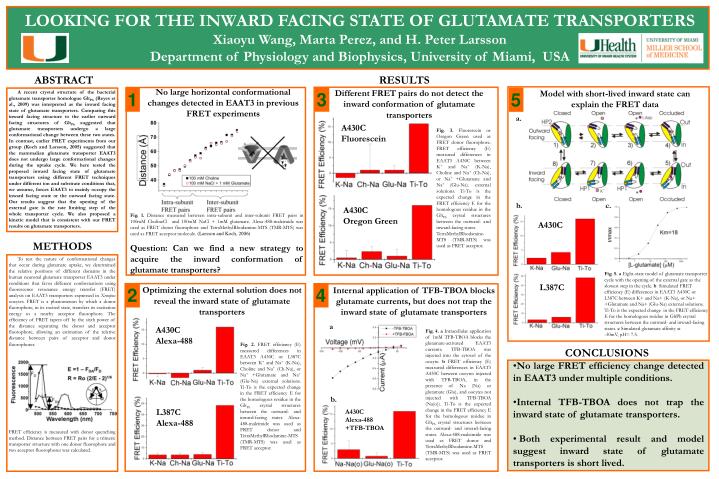
LOOKING FOR THE INWARD FACING STATE OF GLUTAMATE TRANSPORTERS. Xiaoyu Wang, Marta Perez, and H. Peter Larsson Department of Physiology and Biophysics, University of Miami, USA. ABSTRACT. RESULTS. No large horizontal conformational changes detected in EAAT3 in previous FRET experiments.

E N D
LOOKING FOR THE INWARD FACING STATE OF GLUTAMATE TRANSPORTERS Xiaoyu Wang, Marta Perez, and H. Peter Larsson Department of Physiology and Biophysics, University of Miami, USA ABSTRACT RESULTS No large horizontal conformational changes detected in EAAT3 in previous FRET experiments Different FRET pairs do not detect the inward conformation of glutamate transporters Model with short-lived inward state can explain the FRET data 1 3 5 A recent crystal structure of the bacterial glutamate transporter homologue GltPh (Reyes et al., 2009) was interpreted as the inward facing state of glutamate transporters. Comparing this inward facing structure to the earlier outward facing structures of GltPh suggested that glutamate transporters undergo a large conformational change between these two states. In contrast, earlier FRET experiments from our group (Koch and Larsson, 2005) suggested that the mammalian glutamate transporter EAAT3 does not undergo large conformational changes during the uptake cycle. We here tested the proposed inward facing state of glutamate transporters using different FRET techniques under different ion and substrate conditions that, we assume, forces EAAT3 to mainly occupy the inward facing state or the outward facing state. Our results suggest that the opening of the external gate is the rate limiting step of the whole transporter cycle. We also proposed a kinetic model that is consistent with our FRET results on glutamate transporters. 0.4 a. 0.1 A430C Fluorescein Fig. 3. Fluorescein or Oregon Green used as FRET donor fluorophore. FRET efficiency (E) measured differences in EAAT3 A430C between K+ and Na+ (K-Na), Choline and Na+ (Ch-Na), or Na+ +Glutamate and Na+ (Glu-Na). external solutions. Ti-To is the expected change in the FRET efficiency E for the homologous residue in the GltPh crystal structures between the outward- and inward-facing states. TetraMethylRhodamine-MTS (TMR-MTS) was used as FRET acceptor. Zd=4.6eo 0.2 Conductance (nS) Conductance (nS) 0.01 1E-3 0 50 0 50 50 0 50 Voltage (mV) Voltage (mV) 14 1 12 10 b. c. Zd=5.9eo A430C Oregon Green max 8 d Fig. 1. Distance measured between intra-subunit and inter-subunit FRET pairs in 100mM CholineCl and 100mM NaCl + 1mM glutamate. Alexa-488-maleimide was used as FRET donor fluorophore and TetraMethylRhodamine-MTS (TMR-MTS) was used as FRET acceptor molecule. (Larsson and Koch, 2006) Question: Can we find a new strategy to acquire the inward conformation of glutamate transporters? G/G z 6 A430C 4 2 METHODS 0 30 20 10 0 To test the nature of conformational changes that occur during glutamate uptake, we determined the relative positions of different domains in the human neuronal glutamate transporter EAAT3 under conditions that favor different conformations using fluorescence resonance energy transfer (FRET) analysis on EAAT3 transporters expressed in Xenopus oocytes. FRET is a phenomenon by which a donor fluorophore, in its excited state, transfers its excitation energy to a nearby acceptor fluorophore. The efficiency of FRET tapers off by the sixth power of the distance separating the donor and acceptor fluorophore, allowing an estimation of the relative distance between pairs of acceptor and donor fluorophores. FRET efficiency is measured with donor quenching method. Distance between FRET pairs for a trimeric transporter structure with one donor fluorophore and two acceptor fluorophores was calculated. Fig. 5. a Eight-state model of glutamate transporter cycle with the opening of the external gate as the slowest step in the cycle. b Simulated FRET efficiency (E) differences in EAAT3 A430C or L387C between K+ and Na+ (K-Na), or Na+ +Glutamate and Na+ (Glu-Na) external solutions. Ti-To is the expected change in the FRET efficiency E for the homologous residue in GltPh crystal structures between the outward- and inward-facing states. c Simulated glutamate affinity at -30mV, pH= 7.5. L387C 2 4 Optimizing the external solution does not reveal the inward state of glutamate transporters Internal application of TFB-TBOA blocks glutamate currents, but does not trap the inward state of glutamate transporters a A430C Alexa-488 Fig. 4.a Intracellular application of 1mM TFB-TBOA blocks the glutamate-activated EAAT3 currents. TFB-TBOA was injected into the cytosol of the oocyte. bFRET efficiency (E) measured differences in EAAT3 A430C between oocytes injected with TFB-TBOA, in the presence of Na (Na) or glutamate (Glu), and oocytes not injected with TFB-TBOA (Na(o)). Ti-To is the expected change in the FRET efficiency E for the homologous residue in GltPh crystal structures between the outward- and inward-facing states. Alexa-488-maleimide was used as FRET donor and TetraMethylRhodamine-MTS (TMR-MTS) was used as FRET acceptor. Fig. 2. FRET efficiency (E) measured differences in EAAT3 A430C or L387C between K+ and Na+ (K-Na), Choline and Na+ (Ch-Na), or Na+ +Glutamate and Na+ (Glu-Na) external solutions. Ti-To is the expected change in the FRET efficiency E for the homologous residue in the GltPh crystal structures between the outward- and inward-facing states. Alexa-488-maleimide was used as FRET donor and TetraMethylRhodamine-MTS (TMR-MTS) was used as FRET acceptor. CONCLUSIONS • No large FRET efficiency change detected in EAAT3 under multiple conditions. • Internal TFB-TBOA does not trap the inward state of glutamate transporters. • Both experimental result and model suggest inward state of glutamate transporters is short lived. b. L387C Alexa-488 A430C Alexa-488 +TFB-TBOA