Download
1 / 43
440 likes | 687 Views
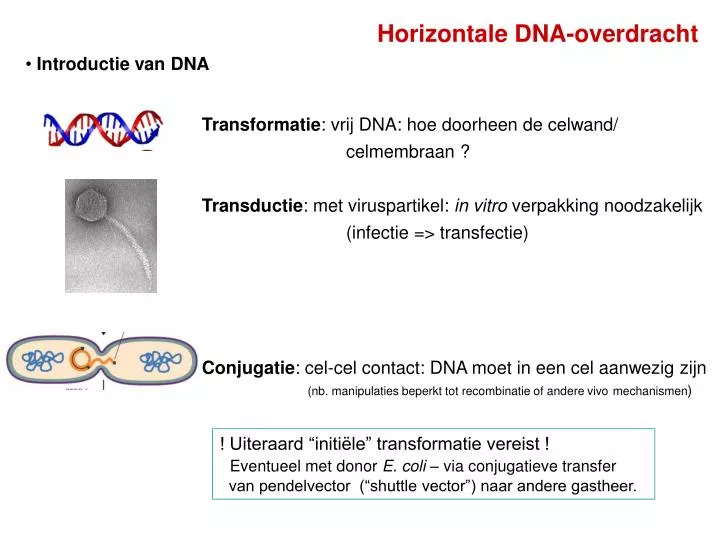
Horizontale DNA-overdracht. Introductie van DNA. Transformatie : vrij DNA: hoe doorheen de celwand/ celmembraan ? Transductie : met viruspartikel: in vitro verpakking noodzakelijk (infectie => transfectie) Conjugatie : cel-cel contact: DNA moet in een cel aanwezig zijn.

E N D
Horizontale DNA-overdracht • Introductie van DNA Transformatie: vrij DNA: hoe doorheen de celwand/ celmembraan ? Transductie: met viruspartikel: in vitro verpakking noodzakelijk (infectie => transfectie) Conjugatie: cel-cel contact: DNA moet in een cel aanwezig zijn (nb. manipulaties beperkt tot recombinatie of andere vivomechanismen) ! Uiteraard “initiële” transformatie vereist ! Eventueel met donor E. coli – via conjugatieve transfer van pendelvector (“shuttle vector”) naar andere gastheer.
Horizontale DNA-overdracht • Illustratie conjugatie
ColE1 : mobiliseerbaar door F F zorgt voor de pili ColE1 heeft nic/bom locus (oriT) en codeert Mob-eiwitten pBR322 : geen mob gebied (eiwitten); wel nic/bom plaats mobiliseerbaar in triparentaal mechanisme F of Ia-type plasmide : pili ColK plasmide : voor de mob-functies (nb. belang van (kennis van) compatibiliteit) pBR327 : heeft geen nic/bom (oriT) meer (idem voor pAT153) niet mobiliseerbaar, tenzij door co-integraatvorming (soms conductie genoemd) transfer m.b.v. mobiliseerbaar plasmide (co-integraat): - recA-afhankelijk : homologe recombinatie met een mobiliseerbaar replicon - recA-onafhankelijk : co-integratie bij transpositie
pSUP plasmiden :(Bielefeld, 1984-1990) • verhoging van efficientie van DNA-overdracht door gebruik van • oriT van incP (RP4, RK2) of incQ (RSF1010) plasmiden • (zie verder voor informatie over deze plasmiden) • - klonering van een Sau3AI oriT-bevattend fragment van RP4 in een • aantal standaardvectoren • pACYC177 => pSUP301 • pACYC184 => pSUP101 • pBR325 => pSUP201, 202 en 203 • - op pSUP202 werd een transposon geplaatst (Tn5) • indien suicide plasmide kan hiermee het transposon • overgedragen worden naar een bacteriestam waarin • het parentale plasmide niet kan repliceren • - in pSUP5011 werd het oriT fragment in de transposonsequentie • geplaatst. • Dit maakt het mogelijk dat na een eerste transfer, waarbij het • transposon (met oriT) op een genetisch element overgedragen • is, dit element te mobiliseren en naar een andere stam verder • te brengen (of terug te brengen naar de oorspronkelijke stam).
nb. het gebruik van de term mob is hier onnauwkeurig (misleidend) en staat door de cis-functie oriT
Integratieve klonering : Vector niet-repliceerbaar (zelfmoordplasmide) Circulair DNA : enkelvoudige homologe recombinatie Gelineariseerd DNA : dubbele homologe recombinatie (uitwisseling) Enkelvoudige recombinatie genereert een herhaling (dus ook instabiliteit) In vitro excisie met restrictie-enzym dat niet in oorspronkelijke construct knipt In vivo uitsplitsing identificeerbaar indien het replicon actief is (vgl. met alleluitwisseling in hoofdstuk over S. cerevisiae) Restrictie-enzyme splitst : - in homoloog gebied : excisie van oorspronkelijk plasmide - buiten het homoloog gebied : herhaling blijft behouden bij excisie => recA- stam vereist voor stabiliteit - in polylinker gebied : herhaling is weg : extensie aan slechts één zijde
Recombinatie tussen homologe sequenties (enkelvoudige cross-over)
Recombinatie tussen homologe sequenties (dubbele cross-over) Integratie via dubbele cross-over in bvb. B. subtilis genoom (Ook methode voor het genereren van deletiemutanten via uitwisseling van actief gen, door gen onderbroken door een resistentiemerker(zie verder) )
Klonering in andere bacteriën (niet E. coli) Waarom? • E. coli • Voordelen : • veel vectoren beschikbaar • eenvoudig transformeerbaar • geschikt voor heel wat toepassingen : o.a. DNA-sequentieanalyse, gerichte mutagenese, • recombinante eiwitexpressie, enz. • Beperkingen: • mist bepaalde ‘pathways’ (bvb. reactiewegen) noodzakelijk voor fenotypische analyse van bepaalde functies van andere bacteriën : • specifieke gastheren nodig – sommige genfuncties vereisen de “eigen” gastheer • post-translationele modificaties van eiwitten • complementatiefuncties, enz.. • Andere gastheer • Vereisten: • DNA moet geïntroduceerd kunnen worden (bvb. door transformatie, conjugatie, enz.) • DNA moet stabiel behouden blijven in de cel (moet functioneren als replicon of in chromosoom ingebouwd worden) • Expressie vanaf plasmide (expressiesignalen: transcriptie/translatie)
Klonering in andere bacteriën (niet E. coli) • Bijkomende vragen: • welke resistenties als merkers te gebruiken ? Gevoeligheid van gastheer • voor antibiotica ? • vectoren: eigen plasmiden ? fagen ? andere ? • restrictiefenomenen (op vreemd DNA) ? • recombinatie ?
Introductie van DNA; horizontale DNA-overdracht Transformatie • “Natuurlijke” mechanismen • meestal transiënt fenomeen • sequentie-afhankelijk of sequentieonafhankelijk • het natuurlijk mechanisme omvat nuclease splitsing van dsDNA, degradatie van • van 1 streng en opname van lineair ssDNA
Conjugatie : - IncPa plasmiden RP4, ~60 kb - IncP plasmiden: transfer van zichzelf en compatibele vectoren - IncP plasmiden kunnen IncQ plasmiden, bvb. RSF1010 mobiliseren - ook transfer mogelijk van E. coli naar Gr+ - relatief groot aantal zelfoverdraagbare plasmiden bij Gr+ Bvb. plasmide pAMb1, origineel Enterococcusfaecalis, ook conjugeerbaar naar E. coli. nb. pAMb1 repliceert volgens een “theta-mechanisme” (zoals ook ColE1) en is stabieler dan heel wat andere plasmiden die volgens een “rolling circle mechanisme” repliceren (hetgeen makkelijker deleties veroorzaakt).
Vectoren afgeleid van IncQ-groep plasmideRSF1010 • multicopy replicon • niet-zelf conjugatief; mobiliseerbaar • resistentie t.o.v. • sulfonamide en streptomycine • unieke restrictieherkennings- • sequenties (RHS) • niet bruikbaar omwille van verlies • van resistentie – interferentie • met expressie • vereiste nieuwe RHS en selectie- • merkers • heel wat derivaten, inclusief pSUP vectoren • RSF1010
Mini-versies van de IncP-groep plasmiden • IncPa plasmide • Vb. RP4 en RK2 ~60 kb, conjugatief en mobiliseerbaar • grote plasmiden • Afgeleide mini-IncP plasmiden ~ 5-7 kb, nog steeds breed spectrum gemeenschappelijke polylinker – lacZ’ regio – blauw/wit selectie oriT = origin of transfer – mobiliseerbaar parDE, parCBA operons: grotere stabiliteit in segregatie trfA : houdt het aantal kopijen laag ; excisie ervan (met NdeI-SfiI) verhoogt het aantal kopijen
Vectoren afgeleid van breed-gastheerspectrum plasmide pSa, Agrobacterium tumefaciens IncW plasmide: pSa grijs : functies nodig voor replicatie paars (donker) : nodig voor zelf-transmissie (tussen SstII en SalI : suppressie van tumorinductie door A. tumefaciens)
Vectoren afgeleid van breed-gastheerspectrum plasmide pSa, Agrobacterium tumefaciens IncW plasmide: pSa - zelfoverdraagbaar naar grote groep Gr- - vector voor “oncogene” A. tumefaciens - 2 regio’s voor conjugatie; 4 kb-regio voor replicatie in diverse gastheren afgeleide vectoren met unieke RHS en insertie-inactivatie van één resistentiemerker door klonering; ook afgeleiden met faag lcos-site : functioneert als cosmidevector. derivaten : niet conjugatief maar mobiliseerbaar door andere conjugatieve plasmiden.
Klonering in andere bacteriën (niet E. coli) Primrose & Twyman hoofdstuk 10 Klonering in gram-positieven • Geen universele vectoren t.g.v. zeer sterke verschillen in GC-gehalte: • Range: <30% tot >70% • onderscheid hoog/laag GC-gehalte organismen • Hoog GC-gehalte, e.g. Streptomyces spp. • Laag GC-gehalte, e.g. Bacillus spp., Clostridium, Staphylococcus, Lactococcus … • Vaak elektroporatie (soms inefficiënt) • Bacillus subtilis – specifieke methoden – Box 10.1
Klonering in andere gram-negatieven (niet E. coli) • In de regel : pendelvector of breed-gastheerspectrum plasmide • Indien klein natuurlijk plasmide beschikbaar optie pendelvector / fusie met • bestaande E. coli vectoren – voordeel indien beschikbare resistentiemerkers • ook in de nieuwe gastheer werken • breed-gastheerspectrum plasmide: • - grootte • - meerdere selectiemerkers (soms) • - unieke restrictieherkenningsposities – bij voorkeur positieve selectie
Behoud van het DNA: • stabiel als plasmide (zie hoger – indien replicatie) • verloren gaan (geschikt voor transposon “delivery”) • integreren in ander replicon, meestal chromosoom (enkele cross-over) • recombinatie tussen homologe sequenties (dubbele cross-over) • Integreren in ander replicon, meestal chromosoom (enkele cross-over) • Optie: selectie voor integratie in geval geen stabiel replicon • pBR322 integratieve vector met neomycine resistentiemerker en gedeelte • amyE (a-amylase gen van B. subtilis) • - selectie op integratie; 1 kopij per cel • - toepassing: naburige sequenties kloneren. • Fig 10.2
Laag GC-gehalte Gr+ Efficiënte transcriptie/translatie • Transcriptie / verschillend van E. coli • RNA polymerase gelijkaardig; evenwel verschillende sigma-factoren • Naast -35 en -10 geconserveerde posities, ook veelal -16 geconserveerd TGTG- • motief weinig voorkomend bij E. coli • Bij mutaties in deze regio, sterk verlies van de promoteractiviteit • Translatie / verschillend van E. coli • B. subtilis ribosomen herkennen enkel eigen mRNA (niet E. coli mRNA) • In tegenstelling tot E. coli ! • Reden ? Missen het S1 ribosomaal eiwit – staat normaal in voor aspecifieke binding • aan RNA en translocatie naar 30S subeenheid (SD alignering en start • translatie vanaf startcodon) – sterker geconserveerde SD-sequentie
Klonering in gram-positieven Laag GC-gehalte Gr+ • Veelal plasmiden van S. aureus gebruikt – ook functioneel in andere Gr+ • pC194, pUB110 – eigen B. subtilis plasmide (pTA1060) • Problemen bij directe klonering in B. subtilis (zie box-info) • pendelvectoren E. coli – B. subtilis • manipulaties in E. coli – plasmide-extractie, multimere plasmiden • te transformeren naar B. subtilis Tabel 10.2 (p.192) • Problemen bij herschikking van grotere inserties- invloed van type replicatie • van plasmide – ‘thèta’ versus ‘rolling-circle’ • ColE1-gebaseerde vectoren via thèta-replicatie • pAMbeta1 = enige ‘thèta’-type replicatie • Hogere stabiliteit / pendelvectoren / mobiliseerbare
Laag GC-gehalte Gr+ Gecontroleerde expressie in gastheren met een laag GC-gehalte Spac systeem in B. subtilis • E. colilacI gen • promoter van faag SPO1 – gekoppeld aan de lac operator • => induceerbaar met IPTG E. coli T7 systeem in B. subtilis • T7 RNA polymerase-gen geïnsereerd in het chromosoom • achter een xylose-induceerbare promoter • Doelwitgen onder controle van T7 promoter op B. subtilis vector Xylose-induceerbare promoter (B. subtilis, Staphylococci, Lactobacillus)
Laag GC-gehalte Gr+ Gecontroleerde expressie in gastheren met een laag GC-gehalte nisA, nis F systemen in Lactococcus lactis • nisA, nisF promoter • induceerbaar met nisine – expressieniveau afhankelijk van nisineconcentratie Andere: Tabel 10.3 Secretie van vreemde eiwitten • Exportmechanismen gelijkaardig aan E. coli • Verschillen in signaalsequentie – meer positief geladen N-terminus + langer • Probevectoren voor selectie van secretiesignaalsequenties zijn gebaseerd op • gesecreteerde reporteractiviteit (S. aureus nuclease) Selectie van secretiesignalen – monitoring van nuclease- activiteit in het medium Prom MCS Nucleasegen
Laag GC-gehalte Gr+ Geninactivatievectoren : studie van genen uit gekende genoomsequenties • studie van genfuncties • rechtstreekse insertionele mutagenese in het chromosoom • pMUTIN vector
Laag GC-gehalte Gr+ pMUTIN vectoren Kenmerken • insertionele mutagenese – geen onafhankelijk replicon in B. subtilis • lacZ reporter voor expressiemonitoring • induceerbare Pspac promoter voor induceerbare expressie Procedure • PCR/doelwitgen – insertie in pMUTIN • transformatie naar B. subtilis • bij integratie: * onderbreking van doelwitgen – eigen promoter wordt • gefusioneerd aan lacZ reporter (geninactivatie – ‘null mutation’) • * stroomafwaartse genen onder controle van Pspac • (conditionele mutatie) • lacZ reporter voor expressiemonitoring • induceerbare Pspac promoter voor induceerbare expressie (IPTG)
Laag GC-gehalte Gr+ pMUTIN vectoren
pSPORTn3 pBR322 en derivaten bevatten nog een inverted repeat element van Tn3 Door het her-invoeren van een tweede element, kan een transponeerbaar element gecreëerd worden. Het transposase (gecodeerd door het tnpA gen) wordt voorzien vanop een additionele (compatibele) vector (bvb. pSC101).
Klonering in andere bacteriën (niet E. coli) Hoog GC-gehalte Gr+ O.a. Streptomyces spp. Algemeen • heel wat spp. maken antibiotica aan – studie synthesewegen • G+C content: ~70-75%, A/T –rijke RHS weinig voorkomend • promotors liggen vaak behoorlijk ver voor de translatiestartplaats • complexe promotersequenties – sommige > 100 bp voor startcodon • (tandemplaatsen voor verschillende sigma-factoren) DNA-opname • Transformatie van protoplasten in aanwezigheid van PEG • 106 – 107 transformanten/µg supercoiled DNA • Elektroporatie na beperkte lysozyme-behandeling (geen protoplastvorming) • Intergenerische conjugatie van mobiliseerbare plasmiden van E. coli naar • Streptomyces (tra genen in trans voorzien; oriT (nic/bom) plaats aanwezig • Antibioticumselectiemerkers vaak moeilijk wegens inherente resistentie van de cel
Hoog GC-gehalte Gr+ Vectoren • RSF1010 repliceert ook in Streptomyces – andere niet • afgeleid van Streptomyces endogene plasmiden of fagen • plasmide: veelal afgeleid van SCP2* aanleiding tot stabiele, laag-kopij aantal • (theta-replicatie) vectoren • vectoren met hoog aantal kopijen (300) afgeleid van het "kleine" plasmide pIJ101 • bijzondere loci voor integratie in chromosoom (o.a. pSAM) f • faag: getemperde faag fC31 ; ook integratieve varianten • BACs waarin 100 kb DNA inbouwbaar is ; pendelvectoren naar E. coli • ook integratievestrategieën (in Streptomyces)
- geïsoleerd in 1955 uit een bodemstaal in New-Mexico (Streptomyces azureus) • - structuur opgelost in 1970 • - slecht oplosbaar in water ; onstabiel in zuur en alkali • actief tegen grampositieven ; door slechte oplosbaarheid geen doorbraak niet in humane geneeskunde • - mogelijks wel ook anti-malaria en antikanker eigenschappen • - (topicaal) veterinair antibioticum (bij mastitis) • - 10 ringen, 11 peptidebindingen, sterk onverzadigd, 17 chirale centra • (toch succesvolle chemische totaalsynthese gerealiseerd) • bindt het GTPase domein in 23S-rRNA ; inhibeert translatie • - toepassing in gentechnologie van Streptomyceten (rRNA methylase als resistentiegen)






















