Download

1 / 25
250 likes | 644 Views
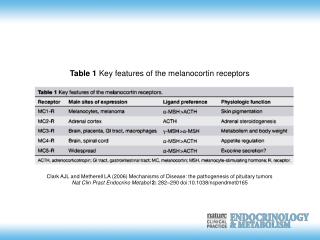
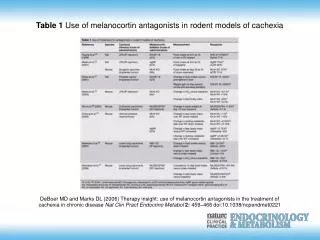
REAKTİF OKSİJEN RADİKALLERİNİN MİTOKONDRİYAL METABOLİZMASI. . ROS yapımı için üç önemli bölge (Kompleks I II III) vardır. Bu ROS’ları yıkan iki majör enzim sistemi MnSOD ve GPX’tir. Mitokondriyal ROS kesinlikle zararlıdır birikmeleri halinde hastalık ve genetik hasar meydana getirir.

E N D
REAKTİF OKSİJEN RADİKALLERİNİN MİTOKONDRİYAL METABOLİZMASI
. ROS yapımı için üç önemli bölge (Kompleks I II III) vardır. Bu ROS’ları yıkan iki majör enzim sistemi MnSOD ve GPX’tir. Mitokondriyal ROS kesinlikle zararlıdır birikmeleri halinde hastalık ve genetik hasar meydana getirir. • Bu maksat ile ROS yapımı ve yıkımı için birden fazla kompleks sistemler vardır. Hangisinin önemli olduğu doku ve metabolik duruma göre değişir.
ROS yapım bölgesinden gelen taşıyıcılar kinetik olarak kontrol edilir. ROS yapımının ölçüldüğü memeli mitokondriyal enzimlerden en az dokuz tanesi tespit edilmiştir. • Bu dokuz enzim, memeli mitokondrisinde aynı anda görülür ve ROS yapım kapasiteleri ekspresyonları doku ve türe göre değişiklik gösterir.
Sitokrom b5 redüktaz: Dış mitokondriyal membrana yakın yerleşimlidir. Bu protein memeli dokularında yaygın olarak bulunur. Okside sitoplazmik NAD(P)H ve indirgenmiş b5 dış membranda bulunur. Ayrıca serbest askorbil radikallerini indirger. Bu askorbatlar rejenerasyonu memeli karaciğerinde saptanır. Bu enzimler şizofreni hastalarında yüksek olarak tespit edilmiştir.
Monoaminoksidaz (MAO): Yine dış membrana yerleşmiş olup aynı anda tüm memeli dokularında ekspresse olur. H2O2 salgılayan biyojenik aminlerin katalizini yapar. Beyin mitokondrisinde monoamin transmitterlerin dönüşümünde esas rolü oynar. H202 yapım gücü diğer mitokondriyal kaynaklardan daha fazladır. MAOlar H202 yapımının iskemik dokuda, yaşlıda, eksojen aminlerin oksidasyonunda majör kaynağıdır. Bazı yazarlar MAO ların artmasıve buna cevap olarak H202 seviyesinin yükselmesinin Parkinson hastalığını arttırdığını öne sürmektedirler.
Dihidroorotat dehidrogenaz (DHOH): Memeli dokularında aynı anda ve mitokondrinin iç zarının dış yüzeyinde yer alır. Pirimidin sentezinde bir basamak olan dihidrooratatı orotata çevirir. Doğal bir elektron alıcısı olan koenzim Q, DHOH yi azaltarak H202 yapar. • Akonitaz: Mitokondriyal matrikste yer alır. Trikarboksilik asit siklusunda yer alan sitratı izositrata çevirir. Bu enzim demir-sülfür bağlı süperoksit tarafından inaktive edilir. İnaktivasyondan sonra izole akonitaza bağlı hidroksil yapımı çoğunlukla salınan Fe’e bağlı görünmektedir.
Alfa gliserofosfatın dehidrogenazı: İç membranın dış yüzeyinde yer alır, farelerin kahverengi yağ dokusu kas, beyin dokusunda yer alır. Gliserol 3 fosfatı dihidrooksaton fosfata çeviren FAD içerikli enzim ile oksidasyonu doğal elektron alıcısı koenzim Q tarafından kullanılır. Bu reaksiyon lipid metabolizması içerir ve NAD dan NADH formu oluşur. Bu enzim ekspresyonu hipertiroid hayvanlarda artar. Fare ve drosofila mitokondrisi gliserol 3 fosfatı okside ederek H202 oluştururlar. Mekanik olarak bu reaksiyonun sonunda drosofilada H202 havuzu enzim başına olması beklenirken bu havuzun %30’ Kompleks I tarafından sağlanır.
Süksinaz dehidrogenaz: İç membranın iç yüzeyinde yer alan bir flavoproteindir. Koenzim Q elektron alıcısı olarak görev yaptığı bir reaksiyonda süksinatı fumarata çevirir. İzole SDH lipozomda ROS yapabilir çoğunlukla FAD yoluyla yapılır. Okside süksinatın submitokondriyal parçası karboksin tarafından inhibe edilir. Bununla birlikte aynı inhibitör ayrıca antimisine bağlı ROS yapımını ve ROS yapımını destekleyen NADH oksidasyonunu süprese eder. Bu iki reaksiyonun SDH dan bağımsız olduğu düşünülmektedir. Bunula birlikte SDH’nın intak solunum zincirinin bir partçası olup olmadığı açık değildir
Alfa-ketoglutarat dehidrogenaz kompleksi: İç membranın matriks yüzüne sıkıca bağlıdır. NAD ın elektron alıcısı olduğu bir reaksiyonda ketoglutarat süksinil CoA’ya okside olur. KGDHC üç ayrı enzimden oluşur: Alfaketoglutarat dehidrogenaz, süksinil transferaz, lipoamid dehidrogenaz. Lipoamid dehidrogenaz bir başka mitokondriyal enzimin pürivat dehidrogenazın parçasıdır. PDHC ve KGDHC süperoksit ve hidrojen peroksit oluşturur. KGDHCden ROS yapımı düşük kullanımlı doğal elektron alıcısı NAD tarafından stimüle edilir.
KOMPLEKS I’DE ROS YAPIMI İzole Kompleks I çalışmalarında ROS’un flavin ve rotenon bölgeleri arasında yapıldığı görüşü vardır. Ayrıca bu bölgenin süperoksit yapım bölgelerinden birisi olduğu düşünülmektedir. Bir başka görüşte Kompleks I’e bağlı ROS yapımının flavine bağlı olduğu yada kompleksin yarısının NAD radikallerine bağlı olduğu düşünülmektedir. Kompleks I’ e bağlı ROS oluşumu ile ilgili üç ayrı deneysel model vardır. Ters elektron transferi (RES), rotenona bağlı ROS yapımı, Normal solunum zincirinde ROS yapımı
RET; başlangıçta submitokondriyal parçalarla aypılan deneysel çalışmalarda bulunmuştur. Mitokondride ROS yapımını gösteren ilk çalışmalardır. RET solunum zincirinde bir reaksiyon seti olup elektron taşıyıcıların redoks potansiyellerine karşı NAD tan Koenzim Q’ya oksijenin alınmasına izin veren reaksiyonlardır. Antimisin A ile tedavi edilen sıkı bir şekilde çift haline getirilmiş submitokondriyal partiküller, süksinatla gelen RET’e NAD eklenmesiyle çok sayıda H202 yapımını gerçekleştirirler. Hem NAD azaltılması hem de ROS yapımı yüksek membran potansiyeline ihtiyaç duyar, çift olmamış elektronlardan korunur
İntak mitokondride süksinatın yada alfa gliserofosfatın oksidasyonu için yüksek membran potansiyeline ihtiyaç duyar. RET ROS yapımını yüksek oranda destekler. ROS oluşumu için yüksek enerjiye ihtiyaç vardır. Transmembran potansiyelinde %10 indirgenme ROS yapımını %90 azaltır. Bununla birlikte enerji harcayan başka bir reaksiyonda da inhibe edilir, örneğin ATP sentezi, Ca uptake, elektron çift yapısının bozulması gibi. RET destekli ROS yapımı matriksin asidifikasyonu ile de süprese edilir, bu durum Kompleks I’in sağladığı katkı bazik pHda gerçekleşir. RET destekli ROS yapımı rotenon tarafından Koenzim Q’dan Kompleks I‘e elektron transferi önlenerek baskılanır.
İkinci deneysel çalışma paradigması rotenona bağlı ROS yapımı; bu durum submitokondriyal partiküllerde ve intak mitokondride NAD bağlı substratlarla piruvat, glutamat eklenmiş malat ile gerçekleşir. Rotenon bağlı ROS yapımı çok yüksek oranda ve derecede redoks taşıyıcı redüksiyonuna ihtiyaç duyar. NAD bağlı substratlar açıkça membran potansiyeli regülasyonuna ihtiyaç duymazlar. Çünkü bu durum Kompleks I tarafından inhibe edilir.
Kompleks I inhibitörlerinin eksikliğinde ROS yapımı NAD bağımlı substratlarca yüksek membran potansiyeli ile stimüle edilir. Bu durum ayrıca Kompleks I’in proksimalindeki proton pompasıda bulunan redoks taşıyıcılarının yüksek derecede redüksiyonu şeklindedir. Membran potansiyel yüksekliğindeki ROS yapım hızının bağlılık durumu RET yerine geçen bir basamak değil termodinmaik olarak daha önemli bir süreçtir.
Q DÖNGÜSÜ VE KOMPLEKS III’TE ROS YAPIMI • . Kompleks III mitokondri solunum zincirinde okside koenzim Q’yu kullanan bir enzim zinciri olup sitokrom c yi elektron alıcısı olarak kullanır. Koenzim Q nun oksidasyonu Q döngüsü olarak adlandırılır.
Spesifik Kompleks III inhibitörleri süperoksit yapımı ve bulunduğu bölgede çok önemli rol oynar. Miksotiazol QH2 nin Qo sitesine bağlanmasını engeller. Antimisin A Qi sitesine ikinci elektronun transferini önler. Kinoldan elektron transferini spesifik olarak ISP (stigmatellin) inhibe eder.
Süperoksite mitokondri membranının geçirgen olduğu son dönem çalışmalarda gösterilmiştir. İç mitokondri zarı süperoksitin kendine yada H202 eklenmiş haline karşı bir bariyer değildir. • Bir raporda Martin Brand matriks akonitozunun süperoksit oluşumunu inhibe ettiğini ve ekstramitokondriyal süperoksit dismutazın bu durumdan koruduğunu göstermiştir. • Ayrıca Kompleks I ve III’ün her ikisininde süperoksit yaptığı, I’ in daha çok mitokondriyal matrikste yaparken III’ün bu işlemi hem matriks hem de sitozolik yüzeyde yaptığını göstermişlerdir.
MnSOD: Mitokondrideki primer ROS yapımına karşı olan ikinci defans sistemidir. Bu ernzim özellikle mitokondri matriksinde yer alır. Süperoksit radikalini dismutasyonla H202’ye çevirir. Mitokondrinin demir- sülfür bileşenlerini süperoksit atağından korur. • Sitokrom C: Süperoksit yakalama kapasitesi olan, mitokondri membranları arasındaki bölgeye yerleşen bir moleküldür. Sitokrom C alternatif olarak mitokondri respiratuvar sisteminde indirgenme yapar. İndirgenmiş sitokrom C doğal elektron alıcısı konumundadır. • Katalaz: ROS’tan üretilen H202 hala daha toksik özellik taşıdığından diğer enzimlerce detoksifiye edilir. Bu enzimlerden birisi de katalazdır. Katalaz, H202’yi O2 ve H20’ya çevirir.
Glutatyon peroksidaz: En çok çalışılan antioksidan enzimdir. GSH yıkımından sonra H202 ve H20 oluşumuna neden olur. Bu seleno enzim memeli dokusunda mitokondri matriksi ve intermembranöz boşlukta yer alır. GPX aktivitesi karaciğer, böbrek, kalpte en yüksektir. Beyin ve iskelet kas mitokondrisinde nispeten düşüktür. Hücresel antioksidan defans sisteminin en önemli parçasıdır. Ayrıca asıl görevi akut yaygın oksidatif strese yanıt vermektir. Mitokondride yavaş yapılan ROS’tan daha öncelikli olarak akut stresi karşılar.
ROS YAPIM VE YIKIM BALANSI • Beyin mitokondrisinin H202’yi yaklaşık 6.5nmol/dak mg protein başına ortadan kaldırdığı gösterilmiştir. Bu yıkım hızı mitokondride ROS yapım hızının 3-12 katı kadar olmuştur. Ancak bu ölçüm dışarıdan verilen H202 nin yıkım hızını gösterir. Endojen yapılanın yıkımını tam olarak yansıtmaz.
Bir çalışmada H202’yi saptayan Horseradish enzimi mitokondri reaksiyonunun başlangıcında ve beşinci dakikasında ölçülmüştür. Bu çalışmadan üç önmeli sonuç çıkarılmıştır: • Sağlam mitokondride en hızlı olarak üretilen ROS’ları bile kendi ROS ortadan kaldırma sistemi ile kontrol edebilir. Fonksiyonel ve yapısal olarak yeterli mitokondri bunu sağlayabilir. • ROS defans sistemleri mitokondrinin değişik yapısal bölümlerinde aktivite gösterir. Matriks içinde ortadan kaldırılan düşük molekül ağırlıklı solüsyonların dönüşümü mitokondrinin net ROS miktarını yansıtır. Glutatyon peroksidaz/ glutatyon redüktaz sistemi en önemli ROS ortadan kaldırma sistemidir. Mitokondriyal glutatyonun alametisin ile ortadan kaldırılması hücre yaşamı ile bağdaşmaz.
Öbür yandan hücre içine giren Ca ROS ortadan kaldırma sistemlerinden GPX1’i inhibe etmekte bu durum da ROS miktarı artışına neden olmaktadır. Bu durum doğrudan stoplazmada glutatyon, NADPH ve membran potansiyeli kaybına neden olur.