Download
1 / 44
440 likes | 690 Views
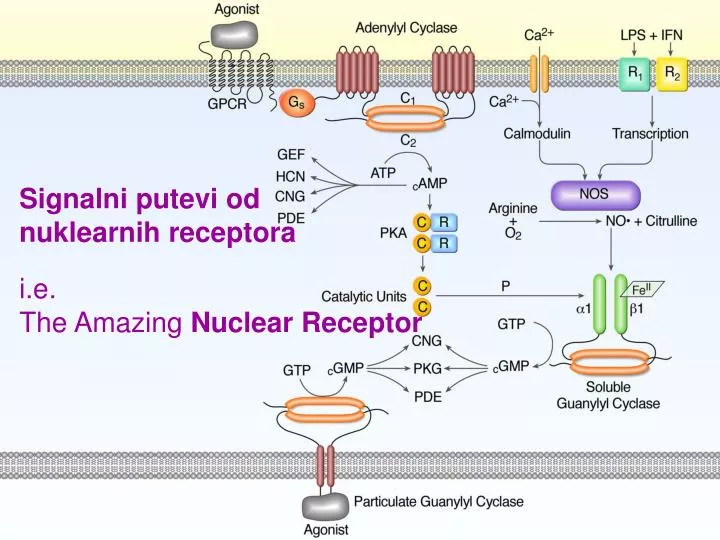
Signalni putevi od nuklearnih receptora. i.e. The Amazing Nuclear Receptor. The Amazing Nuclear Receptor. Ligandi za nuklearne receptore Osnovni principi prenosa signala preko NR Klasifikacija i struktura NR Mehanizmi preko kojih NR regulišu transkripciju Regulacija signalinga preko NR

E N D
Signalni putevi od nuklearnih receptora i.e. The Amazing Nuclear Receptor
The AmazingNuclear Receptor • Ligandi za nuklearne receptore • Osnovni principi prenosa signala preko NR • Klasifikacija i struktura NR • Mehanizmi preko kojih NR regulišu transkripciju • Regulacija signalinga preko NR • Primer: signaling preko NR za steroidne hormone • Membranski receptori za steroidne hormone
SIGNALING PREKO NUKLEARNIH RECEPTORA: • Nuklearni receptori regulišu ekspresiju gena, u zavisnosti od interakcije sa malim lipofilnim molekulama, i na taj način se uključuju u regulaciju velikog broja različitih funkcija ćelija. • To su proteini koji su ligand-zavisni transkripcioni faktori, a nalaze u citoplazmi i/ili nukleusu. • Ligandi, difunduju kroz ćelijsku membranu i vezuju se za receptore u citoplazmi ili u nukleusu. • Kompleks ligand-receptor se vezuje za DNA elemente, u kontrolnim regionima ciljnih gena, i na taj način utiče na transkripciju odredjenog gena.
Ligandi za nuklearne receptore: • Prirodni ligandi za nuklearne receptore su lipofilni hormoni, kao što su steroidni hormoni, tiroidni hormoni, derivati vitamina A i D. Ovi hormoni imaju značajnu ulogu u regulaciji metabolizma, razvoja i diferencijacije, kao i funkcije različitih ćelija u organizmu. Nedavno je utvrdjeno, da i intracelularni lipofilni metaboliti, kao što su prostaglandini, leukotrijeni, masne kiseline, derivati holesterola, žučne kiseline, pa čak i benzoati, mogu da funkcionišu kao ligandi za nulearne receptore, i da na taj način regulišu ekspresiju odgovarajućih gena. • Generalno, ligandi za nuklearne receptore su mali, rigidni hidrofobni molekuli, sa polarnim delovima na jednom kraju, koji im omogućavaju vezivanje za receptore preko Van der Waals-vih i hidrofobnih interakcija, kao i formiranje vodonikovih veza.
Ligandi i HREs strukture za neke nuklearne receptore Skracenice: IR „invertovano ponavljana” (engl. inverted repeated); DR „direktno ponavljana” (engl. direct repeated); ER (engl. everted repeated); NR „bez ponavljanja”. Broj HREs se odnosi na broj parova koji razdvajaju heksamere (pola mesta). NGFI-B (engl. nerve growth factor induced receptor); COUP-TF (engl. Chicken ovalbumin upstream promotor transcription factor); ROR (engl. Retinoic acid releated orphan receptor); , , su subtipovi receptora, kodirani razli;itim genima.
Osnovni principi prenosa signala preko nuklearnih receptora • Nuklearni receptori su ligand-zavisni transkripcioni faktori, koji mogu da funkcionišu kao aktivatori ili represori transkripcije. • Oni se specifično vezuju za HRE, regulatornog regiona ciljnog gena tj. gena čija se transkripcija reguliše. Zavisno od receptora, vezivanje za DNA je povezano sa aktivacijom ili represijom susednih gena. Ipak generalno posmatrano, vezivanje liganda za receptor aktivira transkripciju, a samo u retkim slučajevima dolazi do blokade transkripcije ciljnog gena. • U nekim slučajevima, i receptor koji nije aktiviran ligandom je vezan za DNA , i ima negativan efekat na transkripciju gena. Vezivanje liganda za takav receptor, deblokira aktivaciju transkripcije. • Pored vezivanja sa ligandom, nuklearni receptor može biti aktiviran i angažovanjem drugih mehanizama, kao što su fosforilacija receptora nekom od kinaza, kao i vezivanjem za regulatorne proteine npr. ciklin D1. Drugim rečima, može doći do povezivanja drugih signalnih puteva sa signalingom preko nuklearnih receptora. Signalna transdukcija preko nuklearnih receptora. Nuklearni receptori su ligand-zavisni transkripcioni faktori koji se vezuju za odredjenu DNA sekvencu tzv. HRE. Hormon funkcioniše kao ligand koji reguliše aktivnost receptora. Mnogi receptori funkcionišu kao homo- i heterodimeri. Aktivirani receptor, vezan za DNA inicira transkripciju direktno, ili indirektno preko protein-protein interakcije sa hromatinom i kompleksom proteina koji inicira transkripciju. Strelice pokazuju moguću različitu konfiguraciju HRE. HRE (engl. hormone-responsive element)
Data have implicated nuclear receptors in a number of diseases, including inflammation, osteoporosis, and metabolic syndrome. Researchers use synthetic antagonists for several nuclear receptors to effectively treat asthma, diabetes, atherosclerosis and cancer. Chemicals in the environment can influence receptor signaling pathways directly by functioning as agonists or antagonists of receptors. This interaction may lead to untimely activation of receptors or disruption of receptor signaling. Inversely, nuclear receptors regulate the metabolism of xenobiotics and drugs
The Amazing Nuclear Receptor • Klasifikacija i struktura nuklearnih receptora • Prvi opisani nuklearni receptori su receptori za glukokortikoide, za estradiol i za progesteron. Od tada su otkriveni mnogi drugi, strukturno slični nuklearni receptori, grupisani u veliku superfamiliju sa bar 7 različitih subfamilija. • 48 nuklearnih receptora = identifikovano u humanom genomu • Sisari imaju nuklearne receptore koji su visoko konzervisani „kroz vrste” iako i niži organizmi imaju nuklearne receptore • Svi nuklearni receptori imaju visok stepen homologije na nivou sekvence aminokiselina, što sugeriše da funkcionišu koristeći slične mehanizme. Ipak, još uvek nije poznata funkcija svih nuklearnih receptora, kao i priroda liganda koji se za njih vežu. Moguće je da za neke od ovih receptora tzv. orfanskih receptora ne postoji prirodni ligand i da funkcionišu u ligand-nezavisnom maniru. • “Nuklearni receptori “- Lokalizovani u citoplazmi i/ili u nukleusu i na membrani (steroidni R) • Receptori koji su uvek u nukleusu (R za derivate retinoicne kisene T3 i vitamin D3)
The Amazing Nuclear Receptor Nuclear Receptor Classification Class Endocrine Receptors Adopted Orphan Receptors Orphan Receptors Ligands High affinity, hormonal lipids Low affinity, dietary lipids, xenobiotics Unknown SF-1 LRH-1 DAX-1 SHP TLX RORα,β,γ ERRα,β,γ HNF-4 COUP-TFα,β,γ Examples ERα,β PR AR GR MR RARα,β,γ TRα,β VDR RXRα,β,γ PPARα,β,γ LXRα,β FXR PXR CAR
The Amazing Nuclear Receptor Neki receptori, kao što su PPARs (engl. peroxisome proliferator-activated receptors) i FXR (farnesoidni receptor), mogu da reaguju sa velikim brojem različitih lipofilnih molekula. Zbog toga se smatra da su ovi nuklearni receptori uključeni u regulaciju metabolizma kao i u detoksifikaciju stranih materija. Nuclear Receptors Involved in the Regulation of Drug Metabolism and Transport •PXR–pregnane X receptor (also referred to as SXR) •CAR–constitutive androstane receptor •PPARα–peroxisome proliferator activated receptor α •FXR–farnesoid X receptor •LXR–liver X receptor •VDR–vitamin D receptor
The Amazing Nuclear Receptor Tissue Distribution of Common Nuclear Receptors Hepar PXRLXRVDR FXRPPARαCAR RXRα Gut PXRLXR VDR PPARαCAR RXRα Splien PXRLXRPPARα CAR RXRα
Tipični nuklearni receptor sadrži pet različitih domena, a svaki od njih ima karakterističnu funkciju. To su varijabilni N-terminalni region (A/B), visoko konzervisani domen odgovoran za vezivanje sa DNA (C), linkerni domen (D) koji sadrži signale o nuklearnoj lokalizaciji, ligand-vezujući domen koji je odgovoran i za dimerizaciju (E), i varijabilni C-terminalni domen (F). U A/B regionu, kao i u E i F regionu su nadjeni domeni odgovorni za transaktivaciju (u A/B regionu AF-1 (engl. activation function 1), a u E i F regionu AF-2 (engl. activation function 2)).
HRE = specifična DNA sekvenca za koju se vezuju NR • HREs se obično sastoje od „two consensus hexameric half-sides” (koja je naravno specifična za svaki nuklearni receptor). • Identitet HREs se zasniva na 3 osobine: • Sekvenca baznih parova u heksameru („half-side”) • Broj baznih parova izmedju heksamera („half-sides”) • Relativna orijentacija heksamera • Receptori se za HRE vezuju uglavnom kao dimeri, i to kao heterodimeri ili kao homodimeri. Mali broj nukleranih receptora se veže za HRE kao monomer. U ovom slučaju, HRE sadrži samo jednu kopiju heksamerne sekvence za prepoznavanje, pa se ovi receptori vežu za HRE kao monomeri npr. AGAACA = 1 heksamerna sekvenca za GR • Na osnovu strukture subjedinica nuklearnih receptora, preko kojih se vezuju za DNA, i strukture HRE, mogu se razlikovati četiri kategorije nuklearnih receptora: • Dimeri receptora za steroidne hormone • RXR sadrži heterodimere • Dimerni orfanski receptori • Monomerni “orfanski receptori”
Dimeri receptora za steroidne hormone HREs, za koje se vežu receptori za steroidne hormone, imaju palindromnu strukturu. Npr. glukokortikoidni receptor se kao homodimer, koji se formira u citosolu, veže za invertovano ponovljenu sekvencu, tj. dva heksamera HRE su organizovana kao lik i njegov odraz u ogledalu. Svaka subjedinica dimera, koji je u kompleksu sa DNA, je u kontaktu sa jednim heksamerom HRE = veliki afinitet vezivanja izmedju dimernog receptora i HRE SHRE (genarally found 150-250 bp 5` to the site of transcription initiation) Oestrogen AGGTCANNNTGACCT Dimerni orfanski receptori Orfanski receptori mogu da se vežu za DNA elemente koji sadrže sekvencu koja se direktno ponavlja. U ovom slučaju, receptori se kao homodimeri, vezuju za HRE, koji sadrži dva mesta (heksamera, medjusobno razdvojena različitim brojem baznih parova) koja se direktno ponavljaju. Oligomerna struktura nuklearnih receptora i struktura HREs. A. Vezivanje homodimernog receptora za dvosimetrični palindromni DNA element. (dimer glukokortikoidnog receptora). B. Vezivanje heterodimernog receptora za DNA element koji sadrži ponavljanu sekvencu. C. Vezivanje RXR kao homodimera za HRE koji ima ponovljenu istu sekvencu. D. vezivanje monomernog receptora za HRE sa asimetričnom sekvencom.
TRE (consensus T3 response Element) AGGTCA...AGGTCA TR binds to specific DNA sequences, TR response elements (TREs), as monomers, homodimers, or with RXR in a heterodimer. Since RXR enhances the binding affinity of TR to TRE, TR/RXR heterodimers have been suggested to be major protein complexes that mediate thyroid hormone responses in vivo • RXR sadrži heterodimere • NR za all-trans retinoičnu kiselinu, 9-cis retinoičnu kiselinu, za T3 hormon i za vitamin D3, se vezuju za karakterističnu nesimetričnu sekvencu DNA, koja se ponavlja(engl. direct repeat of recognition sequence) heterodimerne receptore vezane za DNA. • Jedna od subjedinica heterodimera, je uvek RXR (receptor za 9-cis retinoičnu kiselinu) koji se obično veže za 5` kraj HRE. • Heksameri HRE organizovani u maniru “glava ka repu” što zahteva polarnu organizaciju receptornih dimera. • HRE ovog tipa je karakteristično, da razmak izmedju heksamera koji se ponavljaju može biti različit (obično iznosi od 1-5 baznih parova)variranje rastojanja izmedju ponovljenih heksamera može da kreira različite HRE, koje odredjuju koji će se hetero- i homodimeri formirati tj. vezati za sekvencu koju prepoznaju. Monomerni “orfanski receptori” Ovoj grupi receptora pripadaju receptori (npr. RZR; engl. retinoic Z receptor ili NGFI-B; engl. NGF-induced clone B) koji se kao monomeri vežu za asimetricnu sekvencu. Funkcija ovih receptora je još uvek slabo poznata.
DNA-vezujući domen NR • Receptor se za HRE vezuje preko svog DNA-vezujućeg domena, koji je u okviru familije nuklearnih receptora najkonzervisaniji strukturni element (region C u primarnoj strukturi receptora). • DNA-vezujući domen poseduje: • strukturne elemente preko kojih specifično prepoznaje HRE, kao i • elemente koji regulišu dimerizaciju receptora na HRE. • Centralno mesto u DNA-vezujućem domenu zauzimaju dva Zn2Cys4 motiva. Zn-motivi imaju ulogu u pozicioniranju DNA heliksa i nuklearnog receptora, i nisu uključeni u direktnu interakciju sa bazama DNA. N-terminalni Zn-motiv učestvuje u pozicioniranju heliksa preko interakcije sa ostacima šećera i fosfata u kičmi DNA, dok C-terminalni Zn-motiv učestvuje u dimerizaciji i u kontaktu je sa fosfatnim ostacima DNA. Funkcionalni domeni nuklearnog receptora (na primeru glukokortikoidnog receptora). A. AF-1 i AF-2: domeni koji posreduju u stimulaciji transkripcije. B. Model dva Zn++-Cys4 vezujuća motiva na domenu koji se vezuje za DNA. C. Kompleks formiran izmedju dimernih DNA vezujućih domena GR i HRE. Crni krugovi predstavljaju jone Zn++. D. Koncensus sekvenca i konfiguracija HRE.
Za palindromnu sekvencu se vezuju homodimerni receptori, koji se formiraju već u citosolu. Sekvenca i razmak izmedju heksamera u HRE je komplementarna DNA-vezujućim domenima dimernih receptora, kao i njihovom rastojanju. Geometrija dimera receptora odredjuje, kakav razmak izmedju sekvenci prepoznavanja može da se toleriše da bi se formirao kompleks visokog afiniteta.
Ligand-vezujući domen NR • Ligand-vezujući domen (LBD) ima nekoliko značajnih uloga: • vezivanje liganda (agoniste i antagoniste) • odgovoran je za homo- i heterodimerizaciju • formiranje kompleksa sa „heat-shock”proteinima • transaktivaciju i transrepresiju (vezivanje koaktivatora i korepresora). • Vezivanje hormona konformacione promene koje kontrolišu ove uloge a time utiču i na ekspresiju gena.
Elementi transaktivacije NR ligand-nezavisna transaktivacija štiti mesta fosforilacije i mesta interakcije sa koaktivatorima ligand-zavisna transaktivacija Većina nuklearnih receptora poseduje dva strukturna elementa AF-1 i AF-2, koji su uključeni u aktivaciju transkripcije. AF-1 se nalazi u okviru A/B regiona i uključen je u ligand-nezavisnu transaktivaciju. On štiti mesta fosforilacije i mesta interakcije sa koaktivatorima, a njegova uloga još nije u potpunosti razjašnjena. Sekvenca AF-2 domena, koji se nalazi u E regionu, je dobro konzervisana u okviru superfamilije nuklearnih receptora. AF-2 funkcioniše u ligand-zavisnom maniru. AF-2 okružuje heliks H12, koji je deo ligand-vezujućeg domena, kao i druge strukturne elemente E regiona. Kada se ligand veže za receptor, hidrofilna površina H12 heliksa je okrenuta ka spoljašnosti, i obezbedjuje površinu za vezivanje komplementarnih regiona koaktivatora i korepresora.
Mehanizmi preko kojih nuklearni receptori regulišu transkripciju • Funkcije nuklearnih receptora se mogu opisati terminima transkripcione aktivacije i transkripcione represije tj. blokade. Iako je većina nuklearnih receptora uključena u aktivaciju transkripcije (posredovana „pozitivnom” HRE), postoji odredjeni broj receptora koji dovode do blokade transkripcije, kada se za njih veže ligand (posredivana „negativnom” HRE). Takodje, neki nuklearni receptori inhibišu bazalnu transkripciju, tako što su i u odsustvu liganda vezani za „pozitivnu HRE”. • U regulaciju transkripcije preko nuklearnih receptora su uključene višestruke protein-protein interakcije. Pokazano je da nuklearni receptori direktno reaguju sa: • komponentama transkripcionog aparata koji reguliše bazalnu transkripciju (TFIIB i TAF proteini), • koaktivatorima transkripcije, • korepresorima transkripcije, • drugim specifičnim transkripcionim faktorima.
Koaktivatori NR • proteini koji specifično reaguju sa aktiviranim ligand-receptor kompleksom. • Najbrojnija grupa koaktivatora pripada p160 familiji koaktivatora: npr. koaktivator steroidnih receptora 1 (SRC-1, engl. steroid receptor coactivator 1) koncentrisanju HAT aktivnosti u promotornim regionima ciljnih gena. • Druga grupa koaktivatora funkcioniše kao multiproteinski kompleks = npr. TRAP kompleks (TRAP = engl. thyroid hormone receptor activating protein). uspostavljanje veze izmedju receptora vezanog za DNA i holoenzima RNA polimeraze II Reorganizacija hromatina je osnovni korak u aktivaciji transkripcije DNA prekrivene hromatinom. Stoga je acetilacija histona od izuzuzetnog značaja za regulaciju transkripcije. Nuklearni receptor vezan za DNA, može da regrutuje proteinski komplekshiston acetilaznom aktivnošću (HAT).
Korepresori NR T3R i RAR mogu da blokiraju transkripciju u odsustvu liganda. Blokada transkripcije je povezana sa vezivanjem korepresora. Nuklearni korepresori NcoR i SMRT (engl. silencing mediator for retinoic and thyroid hormone receptors), su vezni za ligand-vezujući domen receptora, a od receptora se odvajaju kada se za njega veže odgovarajući ligand Glavni mehanizam preko koga se ostvaruje blokada transkripcije, je angazovanje proteina koji imaju aktivnost histon deacetilaza (HDAC), u promotornom regionu gena. Korepresori nuklearnih receptora, regrutuju proteine sa histon deacetilaznom aktivnosti u promotorni region, i na taj način održavaju tzv. „represovano” stanje hromatina koje sprečava transkripciju.
Model blokade i aktivacije transkripcije preko T3R. U odsustvu T3 hormona, heterodimerni RXR-T3R receptor blokira transkripciju, svojim vezivanjem za TRE (T3 responsive element). Blokada transkripcije se održava, zahvaljujući regrutovanju korepresornog kompleksa proteina, koji ima histon deacetilaznu aktivnost. X = neidentifikovani kofaktor, koji pomaže održavanje bazalnog transkripcionog aparata, vezanog za promotor, u neaktivnom stanju. U prisustvu T3 hormona, korepresori se odvajaju i koaktivatori (SRC/p160 kompleks) se vezuju za heterodimerni receptor. Proteini koji imaju aktivnost histon acetilaze, pomažu u indukciji transkripcije. Takodje, promotor regrutuje i drugi kompleks koaktivatora, DRIP/TRAP, koji je uključen u stabilizaciju vezivanja holoenzima RNA polimeraze.
Regulacija signalinga preko nuklearnog receptora • Regulacija na nivou koncentracije liganda • Regulacija fosforilacijom (delovanje drugih signalnih puteva) –Ser/Thr • Interakcija sa drugim aktivatorima transkripcije • Regulacija preko ubikvitinacije Funkcije domena nuklearnog receptora. A/B, C, E i F domeni nuklearnog receptora, su uključeni u multiplu protein-protein interakciju i na taj način podložni regulatornim uticajima. Kompleksi proteina sa histon deacetilaznom aktivnošću (HDAC) i histon acetilaznom aktivnošću (HAT) se preko korepresora ili koaktivatora vezuju za promotorni region.
From the following article: Principles for modulation of the nuclear receptor superfamily Hinrich Gronemeyer, Jan-Åke Gustafsson & Vincent Laudet Nature Reviews Drug Discovery 3, 950-964(November 2004) Mode of action of nuclear receptors (NRs). After diffusion through the cytoplasmic membrane, the ligand can interact with its cognate receptor where it can exert a 'non-genomic effect' by interacting directly, for example, with kinases (a). The ratio between cytoplasmic and nuclear location can vary between different receptors and is affected by the nature of a ligand. Ligand binding modulates the interaction of the receptor with a plethora of factors. In the absence of ligand, several nuclear receptors are believed to be bound to the regulatory regions of target genes as a corepressor or histone deacetylase (HDAC) complex (b). Histone deacetylation is responsible for the chromatin condensation that accounts for the gene-silencing effect of apo receptors. Ligand binding releases the HDAC complex (c) and results in the recruitment of histone acetyltransferase (HAT) and chromatin-remodelling (CRM) complexes (d). The temporal order and requirement of these complexes can occur in a receptor-, target-gene- and cell-specific manner. In the last step (e), the polymerase II holoenzyme, which comprises the pol II enzyme, TAF (TATA-binding protein-associated factor) and mediator complexes, is recruited and increases the frequency of transcription initiation. B | Schematic illustration of the structural and functional organization of nuclear receptors. The evolutionarily conserved regions C and E are indicated as boxes (green and orange, respectively), and a black bar represents the divergent regions A/B, D and F. Domain functions are depicted above and below the scheme. AD, activation domain; AF1, activation function 1; NLS, nuclear localization signal.
SR • Aporeceptorni kompleks = neaktivni kompleks • Multiproteinski kompleks caperona sa receptorom – Hsp90, Hsp70, Hsp56 • Hsp90 – vezuje se za dimerni R I odrzava R u “unfolded” konformaciji sposobnoj za interakciju sa ligandom • Hsp70 – pomazu pravilno vezivanje Hsp90 sa R • Hsp56 + p23 – ukljuceni u proces aktivacije R Čaperoni u ovom sistemu funkcionišu kao regulatori aktivnosti receptora za steroidne hormone. Njihova funkcija je da fiksiraju receptor u optimalnoj konformaciji koja omogućava da se receptor veže za hormon sa visokim afinitetom = funkcija čaperona je da stabilizuju odredjenu konformaciju, optimalnu za vezivanje hormona. Takodje, neaktivan kompleks nuklearnog receptora vezanog za čaperone, može biti nadjen i u nukleusu. Na primer, estrogeni receptor, se nalazi u nukleusu, u sastavu velikog kompleksa sa Hsp proteinima. Vezivanje estradiola za receptor, omogućava njegovo odvajanje od Hsp, vezivanje za DNA, formiranje dimera receptora, i interakciju sa koaktivatorima i aparatom za transkripciju.
(a) Peroxisome proliferator-activated receptors (PPARs) function as heterodimers with retinoid X receptors (RXRs) and are activated by specific ligands; they then modulate DNA transcription by binding to defined nucleotide sequences (peroxisome proliferator response element, PPRE) in the promoter region of target genes. Several cofactors (coactivators or corepressors) mediate the ability of nuclear receptors to stimulate or repress the transcription process. (b) The N-terminus A /B domain contains a ligand-independent transcriptional activation domain (AF-1), which can be regulated by mitogen-activated protein kinase (MAPK) phosphorylation in and isotypes. The C domain contains two zinc-finger- like motifs that specifically bind the PPRE in the regulatory region of PPAR-responsive genes. The D domain or hinge region allows conformational changes in the molecule. The E /F domain consists of the ligand-binding domain and the ligand-dependent transcriptional activation domain (AF-2). The ligand-binding pocket appears to be quite large in comparison with other nuclear receptors, allowing the PPARs to interact with a broad range of natural and synthetic ligands.
Retinoid-related Orphan Receptors (RORs) The retinoid-related orphan receptors alpha, beta and gamma (RORα, β and γ , also referred to as NR1F1, 2 and 3, respectively) comprise a distinct subfamily of nuclear receptors. Study of ROR-deficient mice has implicated RORs in the regulation of a number of biological processes and revealed potential roles for these proteins in several pathologies. Mice disrupted in RORα expression display an ataxic phenotype caused by severe cerebellar neurodegeneration (see figure 2). In addition, RORα-deficient mice have thin bones, exhibit an altered immune response and develop atherosclerosis on a high fat diet. Recently, studies of mice deficient in the expression of RORγ unveiled several important physiological functions for this orphan receptor. Mice deficient in RORγ lack all lymph nodes and Peyer's patches suggesting that RORγ is essential for the development of secondary lymphoid organs (see figure 2). The deficiency in these secondary lymphoid tissues is due to the absence of lymphoid tissue-inducer (Lti) cells. RORγ-deficient mice also exhibit defects in thymopoiesis. Thymocytes from RORγ knockout mice undergo accelerated apoptosis that is due to a suppression of the expression of Bcl-xL, an anti-apoptotic protein. These observations suggest that RORγ plays a role in controlling homeostasis during thymopoiesis. Recent studies have implicated RORα and RORγ in the regulation of the adaptive immune response. Both RORα- and RORγ-deficient mice exhibit a decreased susceptibility to allergen-induced lung inflammation, which in the case of RORγ-deficient mice may be related, in part, to the absence of proinflammatory TH17 T-helper cells. Additional studies using gene profiling identified important roles for RORα and RORγ in the regulation of genes encoding several Phase I and Phase II metabolic enzymes in liver. These observations suggest that RORα and RORγ receptors have a critical function in the regulation of several metabolic pathways, including those involved in the metabolism of steroids, bile acids and xenobiotics. Therefore, they are important in the control of metabolic homeostasis
ENDOCRINE PHYSIOLOGY AND METABOLISM, September 2008 Rapid signaling by steroid receptors Ellis R. Levin Steroid receptors transcribe genes that lead to important biological processes, including normal organ development and function, tissue differentiation, and promotion of oncogenic transformation. These actions mainly result from nuclear steroid receptor action. However, for 50 years, it has been known that rapid effects of steroid hormones occur and could result from rapid signal transduction. Examples of these effects include stress responses to secreted glucocorticoids, rapid actions of thyroid hormones in the heart, and acute uterine/vaginal responses to injected estrogen. These types of responses have increasingly been attributed to rapid signaling by steroid hormones, upon engaging binding proteins most often at the cell surface of target organs. It is clear that rapid signal transduction serves an integrated role to modify existing proteins, altering their structure and activity, and to modulate gene transcription, often through collaboration with the nuclear pool of steroid receptors. The biological outcomes of steroid hormone actions thus reflect input from various cellular pools, cocoordinating the necessary events that are restrained in temporal and kinetic fashion. Here I describe the current understanding of rapid steroid signaling that is now appreciated to extend to virtually all members of this family of hormones and their receptors.
Ramirez VD Cell Mol Neurobiol. 1996 Apr;16(2):175-98. Membrane receptors for estrogen, progesterone, and testosterone in the rat brain: FANTASY OR REALITY!!! Ramirez VD et al.
Membranski steroidni receptori Steroids. 2002 May;67(6):529-34 Three different mechanisms are currently under discussion as being responsible for these effects: (1) specific interaction with the cytosolic glucocorticoid receptor (cGCR), (2) nonspecific interactions with cellular membranes and (3) specific interactions with membrane-bound glucocorticoid receptors (mGCR).
Hammes, S. R. et al. Endocrine Review 2007;28:726-741 Translocation of Era to the plasma membrane Translocation of ERa to the plasma membrane. Palmitoylation at cysteine 447 of ER promotes the association of the steroid receptor with caveolin 1. The scaffolding domain of caveolin-1 (amino acids 80–100) then facilitates the translocation of caveolin and ER to the caveolae rafts in the membrane. Here, ER associates with a large protein complex, changing in nature depending upon the cell and signal context, to effect G protein activation and downstream signaling to cell biology.
Hammes, S. R. et al. Endocrine Review 2007;28:726-741 Progestins and androgens regulate kinase and G protein signaling Progestins and androgens regulate kinase and G protein signaling. Classical PRs and ARs transactivate the EGFR in response to ligand by signaling through Src (left). Although human PRB can directly activate Src, AR may utilize MNAR as a scaffold. EGFR activation is mediated either intracellularly or through the release of membrane-bound EGF-like proteins (e.g., amphiregulin or HB-EGF). Activated EGFR then promotes MAPK signaling, which subsequently enhances the activity of multiple nuclear transcription factors. MAPK signaling also leads to phosphorylation and nuclear accumulation of PRB. In X. laevis oocytes, androgens promote maturation by signaling through the AR and MNAR to suppress constitutive Gβ and G s activity and lower intracellular cAMP (right). In fish, progestins promote oocyte maturation by signaling through a G i-coupled mPR. Membrane-bound mPRs may also regulate other G i-mediated processes in somatic cells, and endosomal mPRs may regulate calcium flux in response to progestins.
On binding corticosterone, the mineralocorticoid receptor (MR) and glucocorticoid receptor (GR) multimeric protein complex dissociates (panel a). For the effects on gene expression, MRs and GRs, functioning either as homodimers or heterodimers, interact at a glucocorticoid-response element (GRE) and recruit co-repressors or co-activators, whereas GR monomers interact with stress-induced transcription factors (TFs) or other proteins to dampen their transcriptional activity. The rapid responses, which involve steroid-induced conformational changes and reaggregation with other proteins (such as heat-shock proteins (HSPs)), and a putative membrane steroid receptor, remain poorly understood. The effects of steroids on membrane properties are shown in panel b. On binding to corticosterone, MR and GR can regulate the transcription of genes that are involved in controlling the properties of G protein-coupled receptors, ion channels, ionotropic receptors and ion pumps. This leads to changes in the conductance of the plasma membrane. These steroid actions are conditional, slow in onset and long lasting. RE, response element.
Membranski steroidni receptori Mol Endocrinol. 2008 Oct;22(10):2250-9. Consonant with the coupling of ERalpha to pertussis toxin-sensitive G(i/o) proteins, E(2) also activates G protein-activated inwardly rectifying potassium channels, decreasing membrane excitability and slowing the firing of spontaneous APs in hypothalamic GnRH neurons. These findings demonstrate that the dual actions of E(2) on GnRH neuronal membrane excitability, cAMP production, and GnRH secretion are mediated by the dose-dependent activation of ERalpha and ERbeta expressed in hypothalamic GnRH neurons.
Model for the non genomic actions of 1,25(OH)2D. 1,25(OH)2D binds to a putative membrane receptor. This leads to activation of a G protein (GTP displacement of GDP and dissociation of the b and g subunits from the now active a subunit). Ga-GTP activates phospholipase C (PLC) (b or g) to hydrolyze phosphatidyl inositol bis phosphate (PIP2) to inositol tris phosphate (IP3) and diacyl glycerol (DG). IP3 releases calcium from intracellular stores via the IP3 receptor in the endoplasmic reticulum; DG activates protein kinase C (PKC). Both calcium and PKC may regulate the influx of calcium across the plasma membrane through various calcium channels including L-type calcium channels.
Journal of Cerebral Blood Flow & Metabolism 2008 Estrogen as a Neuroprotectant in Stroke, Patricia D Hurn and I Mhairi Macrae
Journal of Cerebral Blood Flow & Metabolism 2008 Estrogen as a Neuroprotectant in Stroke, Patricia D Hurn and I Mhairi Macrae Estrogen's cellular signaling targets and transduction mechanisms. Estrogenic steroids use diverse mechanisms to produce biological effects, and its neuroprotective properties probably are multifactorial. Classic estrogen signaling (H) occurs through a nuclear receptor that acts as a ligand-activated transcription factor (RH), a protein complex that binds to a hormone-responsive element (HRE) within genomic DNA and targets transcription of mRNA and cognate protein expression. The result is enhanced or suppressed efficiency of gene transcription. There are numerous examples of estrogen's ability to upregulate gene expression, including selected members of the nitric oxide synthase (NOS) and bcl-2 families. Recently, estrogen's activation of plasma membrane hormone receptors and downstream intracellular signaling has been postulated (e.g., through mitogen-activated protein kinase). Such events initiate generalized signaling pathways to the nucleus or to membrane-localized effectors. Evidence also indicates that the hormone can act directly at neurotransmitter receptor complexes or at ion channels (e.g., voltage-dependent calcium channels), resulting in altered neuronal current conductance or transcellular ion flux. Crosstalk between nuclear activation and membrane-associated events is likely. Last, estrogen has important non-cell type-specific actions, such as antioxidation and conservation of endogenous free radical scavenging agents.