Download

1 / 34
340 likes | 372 Views
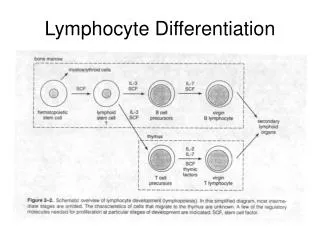
Overview of lymphocyte development. Commitment (متعهد شدن ) Gene rearrangement (نوترکیبی ) Selection (گزینش ). B lymphocyte development. stem cell Pro B cells CD19,CD10, µ heavy chain rearrangement Pre B cells Pre antigen receptor expression immature B cells

E N D
Overview of lymphocyte development Commitment (متعهد شدن ) Gene rearrangement (نوترکیبی ) Selection (گزینش )
B lymphocyte development stem cell Pro B cells CD19,CD10, µ heavy chain rearrangement Pre B cells Pre antigen receptor expression immature B cells IgM BCR expression,independent of antigen and their encounter with it ,receptor editing,celldeath,functional unresponsiveness Mature B cells
نوترکیبی ژنهای گیرنده آنتی ژنی واقعه کلیدی در تکامل لنفوسیت ها بشمار می رود. دو مرحله کنترلی در تولید این گیرنده ها وجود دارد: الف) تولید موفقیت آمیز یکی از زنجیره های پلی پپتیدی ب) تکمیل گیرنده
پیش گیرنده ها و گیرنده های آنتی ژنی سیگنالهاییرا بداخل لنفوسیت های در حال تکامل رها می کنند که برای حیات و بقای آنها مورد نیاز است. درمرحله Immature سلول ها از دو مرحله گزینشی باید عبور کنند. Positive selection and Negative selection در صورتیکه از حیث مرفولوژی و فنوتیپی کامل باشند و گیرنده کاملی در سطح آنها وجود داشته باشد اجازه بلوغ بیشتر پیدا می کنند یعنی انتخاب مثبت می شوندو درغیر اینصورت یا می میرند و یا می بایست دوباره بازآرایی شوند Clonal deletion and Receptor editing و در صورت مفیدبودن سلول ، اجازه می یابند که در مرحله انتخاب منفی مورد آزمایش قرار گیرند . در مرحله گزینش منفی آن دسته از سلولهایی اجازه بلوغ می یابند که گیرنده آنها با تمایل (affinity) ضعیفی با پروتئین های خودی رخورد نماید در غیر اینصورت می میرند.
NK cells این سلولها نقش مهمی در دفاع ذاتی بویژه علیه سلولهای آلوده به ویروس و باکتریهای داخل سلولی بازی می کنند این سلولها 5-15% سلولهای تک هسته ای را در خون و طحال تشکیل می دهند در کبد و رحم حامله هم فراوان هستند. این سلولها در خون بصورت لنفوسیت های بزرگ با سیتوپلاسم پراز گرانول دیده می شوند. این سلولها بر خلاف سلولهای T و B فاقد گیرنده های آنتی ژنی متنوع می باشند. دارای مارکر سطحی CD56 وفاقد مارکر CD3 می باشند دارای مارکر CD16 نیز برای شناسایی سلولهایی که با آنتی بادی IgG پوشانده شده اند می باشند. این سلولها اینترفرون گاما ترشح می کنند که باعث فعال کردن ماکروفاژها برای از بین بردن میکروبهای فاگوسیت شده می گردد.
NK cells مکانیسم کشندگی این سلولها شبیه به سلولهای TCD8 می باشد این سلولها دارای پروتئین های گرانولی می باشند که سلولهای هدف را می کشند. مهمترین پروتئین های توکسیک در گرانولهای سیتوپلاسمی سلولهای NK گرانزیم ها و پرفورین هستند. پرفورین در غشاء سلولهای هدف پلیمریزه شده و منافذی را در غشاء ایجاد میکند و بنابر این موجب تسهیل ورود گرانزیم ها به سیتوزول سلول هدف می گردد. گرانزیم ها آنزیم هایی می باشند که مجموعه ای از سیگنالها را در داخل سلول هدف آغاز می کنندوباعث مرگ سلول هدف از طریق آپپتوزیس می شوند. در ابتدای عفونت ویروسی سلولهای NK زیاد شده و تحت تاثیر IL-12 و IL-15 فعال می شوند و قبل از اینکه سلولهای CTL کاملا فعال شوند سلولهای آلوده را می کشند. این سلولها دارای دونوع گیرنده در سطح خود هستند. الف) گیرنده های مهاری ب) گیرنده های فعال کننده
بلوغ تیموسی اولین سلولی که از مغز استخوان وارد تیموس می شود و درسینوس های زیر کپسولی جامی گیرد Pro-T cell نام دارد این سلولها فاقد TCR وِDouble Neg. می باشند سپس وارد کورتکس شده وبه مرور گیرنده های گاما- دلتا و آلفا - بتا در سطح سلولهای T بیان می شوند. سپس سلولهای T به سلولهای TCD4 و یا TCD8 بالغ می شوند و به مدولا منتقل می شوند و از آنجا تیموس را ترک می کنند.
بلوغ تیموسی محیط تیموس محرک هایی را که برای تکثیر و بلوغ تیموسیت ها لازم است فراهم می کند. در کورتکس ، سلولهای اپیتلیال و در ناحیه بین کورتکس و مدولا ونیز در مدولا سلولهای دندریتیک و ماکروفاژها در مدولا قرار دارند. سلولهای اپیتلیال و دندریتیک ، MHC کلاس I و II را بیان می کنند. سلولهای استرومال تیموس مثل اپیتلیال ها IL-7 را ترشح میکنند که به عنوان فاکتور رشد سلولهای لنفوئیدی عمل می کنند. تیموپویتین حاصل از سلولهای استرومال تیموس نیز در رشد و بلوغ سلولهای T نقش بازی می کنند. گیرنده های Pre-T cell R و TCR سیگنالهایی برای رشد و بلوغ سلولهای T صادر می کنند.
بلوغ تیموسی در کورتکس تیموس سلولهایی که TCR کامل دارند و Double positive هستند مورد گزینش قرار می گیرند درجریان گزینش مثبت ، تیموسیت هایی که به کمپلکس پپتید خودی و MHC متصل شده اند اجازه بلوغ می یابند و تیموسیت هایی که فاقد TCR بوده و یا گیرنده کارآمدی ندارند می میرند. در جریان گزینش منفی آن دسته از سلولهایی که با افینیتی قوی کمپلکس پپتید خودی و MHC را شناسایی می کنند می میرند ودر غیر اینصورت از کورتکس به مدولا مهاجرت می کنند.
بلوغ تیموسی در کورتکس، تیموسیت های Double positive در صورتیکه به کمپلکس پپتید خودی و MHC-I متصل شده اند ژن CD4 آنها خاموش شده و به صورت سلولهای TCD8 باقی می مانند. ودر صورتیکه به کمپلکس پپتید خودی و MHC-II متصل شده باشند ژن CD8 آنها خاموش شده و به سلول های T single positive یعنی TCD4 تبدیل می شوند.
Two types of molecules produced by nonlymphoidthymic cells important for T cell maturation 1)Class-I and II MHC 2)Cytokines and chemokines
سلول Th1 تمايز اين زير گروه از سلول هاي T عمدتا توسط سيتوكاين هاي IL-12 و IFN- γ در پاسخ به ميكروب هايي صورت مي گيرد كه سلول هاي دندريتيك ، ماكروفاژها و سلول هاي NK را فعال مي کنند وجه مشترك همه اين عفونت ها تحريك واكنش هاي ايمني ذاتي است كه منجر به توليد برخي سيتوكاين ها مثل IL-12,IL-18 و اينترفرون هاي نوع يك مي شود. IL-12 قوي ترين محرك تمايز سلول ها ي Th1 بشمار مي رود. در اين رابطه IL-18 نقش سينرژيستيك همراه با IL-12 واينترفرون ها در رابطه با عفونت هاي ويروسي نقش تمايزي براي سلول هاي Th1 بازي مي كنند. موشهاي با نقص در IL-12 و يا IL-12R ، نسبت به عفونت هاي داخل سلولي حساسند.
سلول Th2 تمايز اين زيرگروه از سلول هاي T بوسيله IL-4 در پاسخ به كرم ها و آلرژن ها اتفاق مي افتد. اين گروه از محرك ها موجب تحريك مزمن و مكررسلول هايT مي شوند بنابر اين IL-4 نقش مهمي در راندن سلول هاي T CD4 به سلول هاي Th2 دارد بنظر ميرسد IL-4 اوليه قبل از تمايز سلول هاي Th-2 از ماست سل ها ، بازوفيل ها و ائوزينوفيل ها در واكنش به محرك هاي محيطي و كرم ها سر چشمه مي گيرد.
سلول Th17 تمايزاين سلول ها نتيجه سيتوكاين هاي التهاب زايي است كه در پاسخ به عفونت هاي باكتريايي و قارچي توليد مي شوند. در واقع اين عوامل قارچي از طريق گيرنده Dectin-1 ، سلول هاي دندريتيك را تحريك و آنها را وادار به توليد سيتوكاين هايي مثل IL-1 , IL-6 و IL-23 مي كنند. IL-23 بيش از القاء ، در رابطه با تكثير وابقاء سلول هاي Th-17 اهميت دارد. تمايز سلول هاي TH-17 توسط IL-4 و IFN-γ مهار مي شود بنابر اين پاسخ هاي قوي Th1 و Th2 منجر به سركوب پاسخ هاي Th17 مي شود. جالب اينكه TGF – β كه خود يك سيتوكاين ضد التهابي است در حضور IL-1 و IL-6 موجب تحريك توليد سلول هاي Th-17 مي گردد. يعني TGF – β بطورغير مستقيم مي تواند باعث پيشبرد پاسخ هاي Th17 شود.
Th17 اين سلول ها IL-21 توليد مي كنند كه خود مي تواند با يك مكانيسم تشديد كننده تمايز سلول هاي Th17 را افزايش دهند. طبيعتا در جريان تمايز سلول هاي Th17 نيز فاكتور هاي ترانس كريپشن خاصي نقش دارند كه تحت تاثير سيتوكاين عمل ميكنند . موتاسيون در يكي از اين فاكتور هاي ترانس كريپشن منجر به نقص ايمني كميابي بنام Jobs syndrome مي شود. در اين بيماران آبسه هاي پوستي باكتريايي و قارچي ايجاد مي شود كه نشان دهنده نقص در پاسخ سلول هاي Th17 مي باشد. اين سلول ها در بافت هاي مخاطي بويژه در دستگاه گوارش بفراواني ديده مي شوند شايد بدليل وجود فراوان TGF-β و ساير سايتوكاين ها در اين بافت باشد اين مشاهدات شايد بتواند بيانگر اين واقعيت باشد كه سلولهاي Th17 نقش مهمي در دفاع عليه عفونت ها و التهابات روده اي داشته باشد.
سلول Regulatory T زير گروهي از سلول هاي T CD4 هستند كه پاسخ هاي ايمني را مهار نموده و موجب ابقاء تولرانس محيطي نسبت به خود مي گردد..از ويژگي هاي اين سلول ها ،بروز زنجيره آلفاي گيرنده اينترلوكين 2 (IL_2R chain) يا CD25 و مولكول ترانس كريپشن FoxP3 و ميزان زيادي CTLA-4 ميباشد. نقص در اين سلول ها مي تواند منجر به شيوع بيماريهاي خود ايمن گردد.
Th1 cell Th 1 cell Surface phenotype(αβTCR, CD3, CD4, IL-12R, IFNγR, CXCR3 Effector molecules secreted IFNγ, IL-2, LTα Function Promote protective immunity against intracellular pathogens. By secreting IFNγ they induce activation of macrophages and upregulation of iNOS, leading to the killing of intracellular pathogens such as Leishmania major,Listeria monocytogenes and Mycobacterium spp. Their development is regulated by IL-12.
Th1 Cells T helper type 1 (Th1) cells are a lineage of CD4+ effector T cell that promotes cell-mediated immune responses and is required for host defense against intracellular viral and bacterial pathogens. Th1 cells secrete IFN-gamma, IL-2, IL-10, and TNF-alpha/beta. These cytokines promote macrophage activation, nitric oxide production, and cytotoxic T lymphocyte proliferation, leading to the phagocytosis and destruction of microbial pathogens. In addition to the cytokines secreted by Th1 cells, the expression of specific cell surface receptors, including IL-12 R beta 2, IL-27 R alpha, IFN-gamma R2, CCR5, and CXCR3, can be used to distinguish Th1 cells from other T cell subtypes Th1 cell differentiation and expansion are driven by cytokines that signal through a subset of these receptors, including IL-27, IL-12, and IFN-gamma. Although Th1 cells are critical for the clearance of intracellular pathogens, exaggerated Th1 responses have been found to be associated with autoimmune diseases, including rheumatoid arthritis, multiple sclerosis, inflammatory bowel disease, and type 1 diabetes.
Th2 Cells Th 2 cell Surface phenotype ( αβ TCR, CD3, CD4, IL-4R, IL-33R, CCR4) Effector molecules secreted IL-4, IL-5, IL-13, IL-9 ,IL-10 and IL-17E/IL-25 Function Promote humoral immune responses and host defence against extracellular parasites. However, they can also potentiate allergic responses and asthma. Th2 differentiation occurs in the presence of IL-4 and either IL-2, IL-7, or Thymic Stromal Lymphopoietin (TSLP). IL-4 regulates clonal expansion of Th2 cells, and along with IL-13, promotes B cell production of IgE and alternative macrophage activation. Other cytokines produced by Th2 cells stimulate eosinophil activation and survival (IL-5), or promote mast cell activation (IL-9). In addition to the production of specific cytokines, Excessive Th2-type immune responses have been implicated in the development of chronic allergic inflammation and asthma.
Th9 Cells T helper type 9 (Th9) cells are a distinct subpopulation of CD4+ effector T cell that preferentially secretes high levels of IL-9, CCL17, CCL22, and IL-10 (in mouse). Following activation, naive CD4+ T cells differentiate into Th9 cells in the presence of TGF-beta and IL-4. Under Th9-polarizing conditions, additional cytokines have also been shown to either enhance Th9 differentiation (IL-1 beta, IL-6, IL-21, type I interferons), or promote IL-9 secretion (IL-25, IL-2). Although the signaling molecules and transcription factors involved in Th9 differentiation are still being investigated, it is recognized that Th9 cells are closely related to Th2 cells Unlike Th2 cells, however, Th9 cells do not express IL-4, IL-5, or IL-13. Th9 cells are important for host defense against parasitic helminth infections, but may also have detrimental effects including contributing to the development of chronic allergic inflammation, airway remodeling, and autoimmune disease.
Th 9 cell Th 9 cell Surface phenotype (αβ TCR, CD3, CD4) Effector molecules secreted(IL-9, IL-10) Function Involved in host defence against extracellular parasites, primarily nematodes. Despite their production of anti-inflammatory IL-10, they promote allergic inflammation. Their role in other inflammatory diseases still remains unclear as this subset has only recently been characterized
Th17 Cells T helper type 17 (Th17) cells are involved in the immune response mounted against specific fungi and extracellular bacteria. IL-21 and IL-23 regulate the establishment and clonal expansion of Th17 cells. Cytokines secreted by Th17 cells stimulate chemokine secretion by resident cells, leading to the recruitment of neutrophils and macrophages to sites of inflammation. These cells, in turn, produce additional cytokines and proteases that further exacerbate the immune response. In contrast to mouse Th17 differentiation, Th17 polarization in humans requires IL-1 beta, IL-6, IL-21, and IL-23, but seems to be less dependent upon TGF-beta. One other notable difference is that human Th17 cells secrete IL-26, an IL-10 family cytokine without a murine homologue. Cytokines produced by Th17 cells can have both beneficial and pathogenic effects. While they play a central role in eliminating harmful microbes, persistent secretion of Th17 cytokines promotes chronic inflammation and may be involved in the pathogenesis of inflammatory and autoimmune diseases, including rheumatoid arthritis, multiple sclerosis, and inflammatory bowel disorders
Th 17 cell Th 17 cell Surface phenotype(αβTCR, CD3, CD4, IL-23R, CCR6, IL-1R,CD161 (human only) Effector molecules secreted IL-17A, IL-17F, IL-21, IL-22, CCL20 Function Promote protective immunity against extracellular bacteria and fungi, mainly at mucosal surfaces. Also promote autoimmune and inflammatory diseases. Generated in the presence of TGFβ and IL-6 and/or IL-21 and are maintained by IL-23 and IL-1.
Th 22 It is not antiinflammatory, nor is it necessarily proinflammatory. However,it is consistently described to enable epithelial innate immune responses, which can be detrimental or protective. An example of a detrimental effect is epithelial hyperplasia in psoriasis (PS), which also can be induced by IL-22 in human artificial skin cultures . Furthermore, IL-22 synergizes with IL-17 in the induction of proinflammatory cytokines in human bronchial epithelial cells and colonic myofibroblasts . Examples of protective effects of IL-22 were reported in Gramnegative bacterial pneumonia, in which IL-22 induces the secretion of antimicrobial substances in lung epithelial cells . In addition, IL-22, but not IL-17, protects from hepatitis-induced liver damage by preventing hepatocyte apoptosis . In the skin, IL-22 induces antimicrobial peptides, promotes keratinocyte proliferation, and inhibits differentiation, which suggests a role in remodeling wound healing and in innate defense mechanisms . The role of IL-22 in common skin disorders such as atopic eczema (AE) and allergic contact dermatitis (ACD) is currently not known
Th 22 Th 22 cell Surface phenotype (αβTCR, CD3, CD4, CCR10) Effector molecules secreted IL-22,IL-13, and TNF-alpha Function Identified in inflammatory skin diseases. Their role in host defence remains unclear as this subset has only recently been characterized. Their identity as an independent TH cell subset needs to be confirmed Similar to Th17 cells, Th22 cells express IL-22, CCR4, and CCR6, but in contrast, they also express CCR10 and several fibroblast growth factors (FGFs). In addition, Th22 cells do not express IL-17, CCL20, IL-23 R, CD161 (Th17 markers), IL-4 (Th2 marker), or IFN-gamma (Th1 marker). Collectively, these characteristics distinguish Th22 cells as a novel T helper cell lineage that is distinct from the Th17, Th2, and Th1 subtypes. Activated naive CD4+ T cells differentiate into Th22 cells in the presence of IL-6 and TNF-alpha. Expression of the CCR4 and CCR10 skin-homing receptors on Th22 cells suggests that these cells are likely recruited to the skin, where they may contribute to host defense against microbial pathogens, and promote tissue repair or remodeling. Multiple studies indicate that Th22 cells may also be involved in the pathogenesis of inflammatory skin disorders such as psoriasis, atopic eczema, and allergic contact dermatitis
T Follicular Helper (Tfh) Cells Follicular helper T (Tfh) cells are a distinct subset of CD4+ helper T (Th) cells that regulate the development of antigen-specific B cell immunity.Tfh cell Surface phenotype αβ TCR, CD3, CD4, CXCR5) Upon exposure to a foreign antigen, Tfh cells help B cells generate antibody-producing plasma cells and long-lived memory B cells. Tfh cells are identified by elevated expression levels of multiple surface proteins and Bcl-6, as well as enhanced IL-21 secretion. The high expression levels of these proteins correlate with Tfh cells' enhanced capacity to facilitate antibody production. In the T cell zone of secondary lymphoid tissue, antigen-presenting dendritic cells activate naïve CD4+ T cells to produce IL-21. IL-6 and the autocrine action of IL-21 induce the activated CD4+ T cells to express Bcl-6, the master transcription factor that controls Tfh cell differentiation. Bcl-6, a transcriptional repressor, suppresses the expression of factors that mediate the differentiation of Th1, Th2 and Th17 cells, and clusters of miRNAs that negatively regulate molecules involved in Tfh functioning, such as the CXCR5. Increased expression of CXCR5 helps Tfh cells localize to B cell follicles, where they interact with germinal-centre B cells and secrete a variety of cytokines that stimulate B cells to generate antibody-producing plasma cells and memory B cells.
Regulatory T Cells (Tregs) The primary function of regulatory T cells, also known as suppressor T cells, is to maintain immune homeostasis. This involves suppression of successful immune responses and control of self versus non-self recognition. Failure of the latter results in autoimmune destruction of host cells and tissue. Like other T cells, regulatory T cells mature in the thymus where they are characterized by the variable expression of CD8, CD4, CD25 and FoxP3.. taking advantage of the immunosuppressive potential of regulatory T cells is an important area for advancement in the fields of autoimmune disease and organ transplantation.
Natural Killer T (NKT) Cells Natural killer T (NKT) cells are a specialized population of T cells that express a semi-invariant T cell receptor (TCR alpha beta) and surface antigens typically associated with natural killer cells. The TCR on NKT cells is unique in that it recognizes glycolipid antigens presented by the MHC I-like molecule CD1. Most NKT cells, known as type I NKT cells, express an invariant TCR alpha chain and one of a small number of TCR beta chains. NKT cells can have either protective or deleterious effects due to their abilities to produce cytokines that promote either inflammation or immune tolerance. As a result, they contribute to antibacterial and antiviral immune responses, promote tumor-related immunosurveillance or immunosuppression, and inhibit or promote the development of autoimmune diseases. Like natural killer cells, NKT cells can also induce perforin-, Fas-, and TNF-related cytotoxicity, but this is generally not thought to be their primary function.
NK T cells جمعیت کوچکی از سلولهای T برخی از مارکرهای سلولهای NK مثل CD56رانشان می دهند و آنها را NK T cell می نامند. این سلولها دارای TCR- با تنوع محدود می باشند.پس از فعال شدن سیتوکاین های IL-4,IFN- تولید می کنند و میتوانند به سلولهای B مارژینال در تولید آنتی بادی علیه آنتی ژنهای لیپیدی کمک کنند. قادر به شناسایی آنتی ژنهای لیپیدی همراه بامولکولهای CD1 می باشند. این سلولها ممکن است واسط ایمنی ذاتی علیه برخی از پاتوژنها مثل میکوباکتریوم ها باشند.
gamma delta T Cells Gamma delta T cells represent a small fraction (1 - 5 %) of the overall T cell population but are enriched (more than 50 % of the T cell population) in epithelial cell-rich compartments like skin, the digestive tract, and reproductive organ mucosa. Gamma delta T cells are a subset of T cells defined by the genetic composition of their T Cell Receptor (TCR). All T cells are derived from common progenitor thymocytes, and while the majority of T cells express TCR chain heterodimers encoded by the alpha and beta gene loci, gamma delta T cells express TCR chains encoded by the gamma and delta gene loci. Like helper T cells, gamma delta T cells secrete particular effector cytokines in a subtype-and context-specific manner, however, unlike alpha beta T cells, most delta gamma T cells lack CD4 and CD8 and share a number of markers associated with natural killer cells or antigen-presenting cells such as Fc gamma RIII/CD16 and Toll-like receptors.
(CD8+ cytotoxic T lymphocytes (CTLs are a subset of T cells that express an alpha beta T cell receptor (TCR) and are responsible for the direct killing of infected, damaged, and dysfunctional cells, including tumor cells. CD8 expression and IFN-gamma production are the most commonly used markers for CTL identification. Additionally, Perforin and Granzyme B, which are required for CTL-mediated cell death, are commonly utilized as secondary markers It has been reported that CTLs also produce IL-10, an anti-inflammatory cytokine, at the peak of infection to prevent excessive tissue damage. Importantly, CTLs appear to be critical for cancer prevention as Perforin suppresses lymphoma in mice and mutations in Perforin are associated with lymphoma in humans.