Download
1 / 17
200 likes | 458 Views
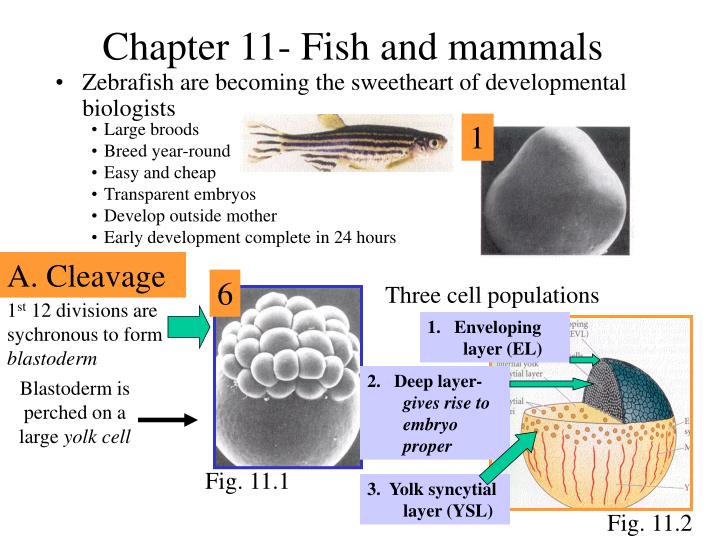
Chapter 11- Fish and mammals. Large broods Breed year-round Easy and cheap Transparent embryos Develop outside mother Early development complete in 24 hours. 1. Zebrafish are becoming the sweetheart of developmental biologists. A. Cleavage. 6. Three cell populations.

E N D
Chapter 11- Fish and mammals • Large broods • Breed year-round • Easy and cheap • Transparent embryos • Develop outside mother • Early development complete in 24 hours 1 • Zebrafish are becoming the sweetheart of developmental biologists A. Cleavage 6 Three cell populations 1st 12 divisions are sychronous to form blastoderm 1. Enveloping layer (EL) 2. Deep layer- gives rise to embryo proper Blastoderm is perched on a large yolk cell Fig. 11.1 3. Yolk syncytial layer (YSL) Fig. 11.2
B. Gastrulation Recall Epiboy from Ch 9 Deep cells migrate to outside then encase entire yolk Epiboly Movement not by crawling, but by YSL cells expansion and pulling EL cells along Embryonic shield epiblast 1. Enveloping layer (EL) hypoblast 2. Deep cells Fig. 11.3 2. YSL cells YSL 6 hrs post-fertilization • A hypoblast is formed either by involution of superficial cells or by ingress • These combine with superficial epiblast cells to form the embryonic shield (function equivalent of the dorsal lip in amphibians)
B. Gastrulation (cont.) The hypoblast cells extend in both directions to form the notochord precursor Animal Head Ventral Dorsal Ectoderm Trunk Tail Fig. 11.3 Vegetal Mesoderm Endoderm Fig. 11.2 -A zebrafish fate map
Chordino Embryonic shield C. Axis formation 1. Dorsal ventral axis- As with the amphibian dorsal lip (Organizer), the embryonic shield: • Establishes the dorsal-ventral axis • Converts lateral/ventral medoderm to dorsal mesoderm (notochord) • Convert ectoderm to neural rather than epidermal B-catenin 2. Forms the notochord precursor samois • 3. Secretes proteins to inhibit BMP from inducing ectoderm to become epidermis • This inhibiting molecule is called Chordino • If mutate chordino, no neural tube is formed Fig. 11.6 goosecoid BMP2 BMP inhibitors e.g. Chordino • 4. Acquires its function from B-catenin accumulation in nearby cells • B-catenin accumulates in YSL cells • Goosecoid is activated
C. Axis formation (cont.) 2. Anterior-posterior axis- In amphibians , the anterior-posterior axis is formed during oogenesis This axis is stabilized during gastrulation by two signaling centers Anterior neural inducing signal (from ectoderm cells) Fig. 11.6 Posterior neural-inducing signal ( from mesoderm cells) 3. Left-right axis - Not much known, but involves TGF-b family signaling molecules
Mammalian Development Tough to study!! • 100um diamater (1/1000th volume of frog egg! • Few in number (<10/female) • Develops within mother • Cleavage events take 12-24 hours each • Development occurs en route to uterus 3. Cleavage during migration down oviduct 2. fertilization 4. Implant in uterus 1. Egg released from ovary Fig. 11.20
Amphibians Mammals A. Cleavage Mammalian Development Distinctions of mammalian cleavage • Slow- 12-24 hrs per cleavage • 2nd cleavage is rotational • Marked asychrony in early cell division • Cleavage at 2nd division requires newly made proteins from zygote Fig. 11.21-rotational cleavage in mammals 5. Compaction (marked cell huddling) occurs at 8 cell stage compaction Fig. 11.23- compaction at 8 cell stage (day 4 in humans)
A. Cleavage (cont.) 16 cell embryo is termed “morula” • external cells will become trophoblast, which will become the placenta • Internal cells will become inner cell mass (ICM), or the embryo proper This marks 1st differentiation event in mammalian development At 64 cell stage, an internal cavity appears and the embryo is termed a blastocyst, ready for implantation onto uterus wall • The Zona pellucida (recall ch. 7) must be shed in order to implant • Blastocyst lyses a small hole in zona using the enzyme strypsin Note- attachment of embryo to oviduct wall is called a tubal pregnancy
B. Gastrulation Similar to reptiles and birds • Mammalian embryo relies on mother for nutrients, not yolk • Thus, the embryo must have a specialized organ to accept nutrients- called the chorion • The chorion induces uterine cells to become a decidua (rich in blood vessels) Epiblasts form amnionic cavity epiblasts Hypoblasts (from ICM) line the blastoceol- these give rise to extraembryonic endoderm hypoblasts blastocoel Fig. 11.28- Day 15 human embryo
B. Gastrulation (cont.) Mammalian mesoderm and endoderm cells arise from epiblasts that migrate through primitive streak E-cadherin attachment is mechanism Direction of migration Henson’s Node Primitive streak Fig. 11.28- Day 16 in human Fig.11.11- Chick gastrulation- similar to mammalian Those cells that migrate through the Henson’s node will become the notochord
B. Gastrulation (cont.) Extraembryonic membrane Formation Trophoblast cells (originally termed “cytotrophoblast) gives rise to multinucleated syncytiotrophoblasts Uterine wall • These syncytiotrophoblasts: • secrete proteolytic enzyme to invade uterine wall • Digest uterine tissue • Mothers blood vessels contact the syncytiotrophoblast cells • Embryo produces its own blood vessels Fig. 11.27-Blastocyst invading uterus Embryo’s blood vessels Chorion Villi Blood vessels feed embryo, but blood cells do not mix Embryo chorion Mother’s Placenta Fig. 11.31 Mothers blood vessels
C. Anterior-posterior axis formation • Two signaling centers • Anterior visceral endoderm (AVE) • Node (Organizer) These work together to form forebrain Fig. 11.34 These are on opposite sides of a “cup” structure Node produces Chordin and Noggin Knock-out of one of these results no forebrain AVE produces Lim-1 and Otx-1
C. Anterior-posterior axis formation The Hox genes specify anterior-posterior polarity These are homologous to homeotic gene complex (Hom-C) of drosophila Recall that the Hom-C genes are arranged in the same order as their expression pattern on anterior-posterior axis Mammalian counterparts are clustered on 4 chromosomes Equivalent genes (Hoxb-4 and hoxd-4) are called a paralogous group
C. Anterior-posterior axis formation (cont.) Fig. 11.36- Hox genes are organized in a linear sequences that concurs with posterior to anterior structures This is referred to as the hox code
Incus Stapes Hox gene rules 1. Different sets of Hox genes are required for specification of any region of the anterior-posterior axis Hoxa-2 KO- stapes missing, duplicate incus Hoxa-3 KO- thymus, neck cartilage malformed 2. Different members of a paralogous group may specify different organ subsets in a given region Hoxd-3 KO deformed atlas (1st vertebra) Example Hoxa-3/Hoxd-3 double KO- atlas and neck cartilage nearly absent 3. A hox gene KO causes defects in the anterior-most region of that gene’s expression
Retinoic Acid has a profound effect on development Recall amphibian development (Ch. 10) Structure of retinoic acid (not in textbook) Fig. 10.41 Hox gene RA Retinoic acid bind a receptor, then the complex binds promoter of a hox gene Retinoic acid activates mammalian hox genes Retinoic acid is likely produced in the node, and perhaps more time spent in the node dictates more posterior specification Lacks all distal vertebra Wild-type mouse embryo RA-treated mouse embryo
D. Dorsal-ventral axis formation Dorsal axis forms from ICM cells near trophoblast Inner cell mass (ICM) Ventral axis forms from ICM cells near blastcoel Trophoblast Blastocoel Fig. 11.32 Fig. 11.42 E. Left-right axis formation Note that mammals are asymmetrical Two levels of regulation- 1. Global- a inv gene defect results in all organs on the wrong side 2. Organ-specific- an iv gene defect causes the axis of an organ to change Organs are located in specific locations