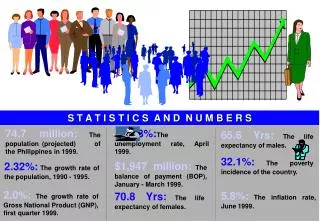
Download

1 / 29
621 likes | 1.5k Views
CHƯƠNG 4. QUÁ TRÌNH TÁI BẢN VÀ SỬA CHỮA ADN. I. Quá trình tái bản DNA ở Prokaryota. Đặc điểm Theo cơ chế bán bảo tồn : pt ADN mới gồm 1 mạch cũ và 1 mạch được tổng hợp mới. Thành phần tgia : AND khuôn dNTP: 8 loại ADN helicaza: cắt liên kết hydro giữa 2 mạch đơn

E N D
CHƯƠNG 4. QUÁ TRÌNH TÁI BẢN VÀ SỬA CHỮA ADN
I. Quá trình tái bản DNA ở Prokaryota Đặc điểm Theo cơ chế bán bảo tồn: pt ADN mới gồm 1 mạch cũ và 1 mạch được tổng hợp mới. Thành phần tgia: AND khuôn dNTP: 8 loại ADN helicaza: cắt liên kết hydro giữa 2 mạch đơn Pr SSB: giữ cho sợi đơn ở dạng thẳng ADN polymeraza (I +III): tổng hợp và sửa chữa AND ADN ligaza: nối hoàn thiện ADN ADN topoisomeraza: mở xoắn ptu AND ARN polymeaza, primeaza: tổng hợp đoạn ARN mồi.
Bảng 5 - 1. Các protein và gen liên quan đến tái bản DNA ở E.coli
I. Quá trình tái bản DNA ở Prokaryota2. Diễn biến: 3gđ 2.1. Giai đoạn mở xoắn: Enzyme topoisomeraza tham gia mở xoắn, 2 loại: Topoisomeraza I: tháo xoắn dạng ADN siêu xoắn -> gắn vào ADN -> cắt một trong 2 sợi -> tạo sợi ADN tháo xoắn sau đó nối chỗ đứt lại Topoisomeraza II: cắt cả 2 mạch của phân tử DNA (gyraza của E.coli) Liên kết Hydro giữa 2 sợi đơn bị bẻ gãy nhờ E helicaza. Hai sợi tách nhau tạo chạc tái bản có cấu trúc chữ Y. Protein SSB gắn vào chuỗi đơn DNA ổn định trạng thái sợi đơn DNA (tránh chuỗi xoắn trở lại).
I. Quá trình tái bản DNA ở Prokaryota2. Diễn biến (tiếp) 2.1. Giai đoạn mở xoắn (tiếp) QT tái bản bđ từ 1 điểm (điểm khởi đầu sao chép –origin) sau đó lan ra theo 2 hướng đến khi đụng vào nhau ở điểm nối đối diện, tạo thành 2 NST vòng xoắn. Vị trí đứt khởi đầu tái bản thường nằm ở những vùng ADN có chứa nhiều trình tự AT, dài từ 100 - 200bp. Vi khuẩn chỉ có một đơn tái bản (replicon).
I. Quá trình tái bản DNA ở Prokaryota2. Diễn biến (tiếp) 2.2. Giai đoạn kéo dài Tổng hợp đoạn RNA mồi: nhờ ARN pol và primeaza: tổng hợp đoạn ARN ngắn (8 – 12 Nu) Enzym ADN pol III tổng hợp mạch bổ sung từ đầu 3’OH tự do của mồi ARN. Mạch khuôn được sd đến đâu các Pr SSB được giải phóng ra đến đó. Chiều tổng hợp DNA từ 5’->3’ -> mạch đơn mới được tông hợp theo 2 hướng ngược nhau: Trên mạch khuôn hướng 3’-5’: mạch đơn mới được tổng hợp theo chiều cùng hướng với hướng tháo xoắn, mạch này được tổng hợp liên tục -> mạch tiến. Trên mạch khuôn 5’-3’: mạch đơn mới được tổng hợp theo hướng ngược với hướng tháo xoắn, xảy ra không liên tục mà thành những đoạn ngắn - đoạn Okazaki (1000 - 2000 bp) -> mạch chậm hay lùi.
I. Quá trình tái bản DNA ở Prokaryota2. Diễn biến (tiếp) 2.3. Giai đoạn hoàn chỉnh sợi mới tổng hợp Sau khi tái bản kết thúc, các mồi RNA bị phân rã bởi hoạt tính 5’ – 3’ exonucleaza của DNA pol I -> tạo lỗ hổng -> được lấp đầy nhờ chính enzyme DNA pol I Enzyme ligaza sẽ nối tất cả các chỗ gián đoạn trên mạch mới. -> Kết quả: tạo 2 chuỗi DNA xoắn kép mới. Tại mỗi chuỗi mới có 1 mạch DNA của DNA mẹ ban đầu. Mạch còn lại mới được tổng hợp thêm. Các enzim tác động rời khỏi phân tử DNA ra môi trường .
II. Quá trình tái bản ở Sinh vật Eukaryota1. Đặc điểm • Cơ chế tái bản ở sinh vật nhân chuẩn cũng giống như sinh vật tiền nhân theo cơ chế bán bảo thủ.Tuy nhiên có một số điểm khác biệt: • Có nhiều loại ADN polymeraza tham gia vào QT sao chép: • Polymerase /primase: tổng hợp mồi RNA cho mạch chậm , không có khả năng sửa sai (exonuclease) do đó không phải là thành phần duy nhất tham gia vào quá trình tái bản. • Polymerase : cn giống DNA polymerase I ở sinh vật tiền nhân (vừa tổng hợp vừa sửa chữa và hoàn chỉnh sợi đơn DNA sau khi mồi RNA được loại bỏ). • Polymerase được tìm thấy ở ty thể có chức năng chưa rõ. • Polymerase : có cn gần với DNA polymerase III ở sinh vật tiền nhân. • Polymerase mới được phát hiện gần đây có vai trò chưa rõ. • Có sự tham gia của nhiều Pr chuyên biệt (VD: Nhân tố kết gắn nhiễm sắc CAF-1) • Đoạn khởi đầu tái bản dài (từ hàng chục ngàn đến hàng trăm ngàn bp - giầu AT); có nhiều điểm khởi đầu trên một nhiễm sắc thể và trong genome.
II. Quá trình tái bản ở Eukaryota 2. Mô hình tái bản: • Đầu tiên, phức hợp nhận biết vùng khởi đầu tái bản (ORC) bọc lấy những điểm khởi đầu, dưới tác dụng của enzyme topoisomerase và nhân tố tái bản (RF-A) làm DNA được tháo xoắn. • Trên mạch chậm, polymerase /primase tương tác với RF-A để tổng hợp lên các mồi RNA dài độ 10 nucleotide, mồi này sau đó được nối dài thêm 20 dN nữa nhờ enzyme polymerase kết hợp với nhân tố sao chép C (RF-C). Khi đó phức hợp PCNA-ATP đã chặn polymerase lại, để cho phép enzyme polymerase gắn vào chuỗi và tổng hợp đoạn Okazaki. • Enzyme polymerase sau khi được giải phóng sẽ chuyển đến mạch đối diện và tổng hợp liên tục mạch tiến. • Các phtử DNA ngay sau khi được sao chép đã tổ chức lại ngay thành các nucleosome.
II. Quá trình tái bản ở Eukaryota 3. Kết thúc tổng hợp tại đầu telomere của mỗi NST • Ở mạch tiến: sợi đơn mới được kéo dài từ điểm khởi đầu đến hết đầu mút NST • Mạch lùi: còn một đọan kết thúc trống chưa chạm đến đầu mút 3’ -> Enzyme telomerase gắn thêm những đoạn 3’ AACCCCAAC 5’ vào đầu 3’ của DNA telomere. • Enzyme telomerase RNA di chuyển dọc theo phân tử DNA (sang phải) -> đầu 3’ tiếp tục được kéo dài. • Enzyme primase tạo mồi 3’AAC 5’; DNA polymerase sử dụng đầu 3’ rất dài này làm nguyên bản để lấp đầy đầu cuối cho mạch đơn DNA kia. -> Mồi bị loại bỏ và ligase sẽ nối chỗ trống lại. Kết quả giữ nguyên được chiều dài của NST qua mỗi lần tái bản.
II. Quá trình tái bản ở Eukaryota 4. Cơ chế bảo vệ đầu telomere của NST • Để tránh mất vật liệu di truyền sau mỗi lần tái bản, telomere cần phải kết hợp với một số protein để hình thành lên mũ bảo vệ. • Cấu tạo của mũ bảo vệ telomere: gồm 3 protein là TRF1, TRF2 và WRN, chúng liên kết với nhau rồi bọc lấy những trình tự lặp lại ở telomere, ẩn dấu đầu cheo 3’ (dài đến 100 bp). • Vai trò của mũ bvệ ở telomere: • Ngăn cản không cho enzyme deoxyribonuclease phân giải đầu mut cua phân tử DNA • Ngăn cản không cho các nhiễm sắc thể trong nhân dính vào nhau. • Tạo sự ổn định trong quá trình tái bản DNA ở phần cuối NST. • Nếu không có mũ bảo vệ telomere thì đầu cuối sợi NST sẽ bị gẫy làm NST không ổn định và dần ngắn lại qua mỗi lần tái bản, là nguyên nhân gây ra bệnh ung thư và nhiều bệnh khác liên quan đến quá trình già hoá.
III. Sửa chữa và bảo vệ DNA • Là hoạt động khắc phục sai hỏng trên DNA, do các tác nhân gây đột biến vật lý và hóa học của môi trường trong quá trình tái bản. • DNA là phân tử duy nhất trong tế bào có khả năng được sửa chữa khi bị biến đổi hay phá hỏng ổn định di truyền . • Sai sót thường gặp: • Khoảng 5000base/ ngày bị mất (khử nhóm amin) • Khoảng 1000base/genome/ngày biến đổi CU • Tuy nhiên tần số đột biến về trật tự aa thấp, tỷ lệ đột biến 10-9/thế hệ do cơ chế sửa sai DNA rất nghiêm ngặt và được thực hiện thường xuyên. • Quá trình sửa sai trên DNA liên quan chặt chẽ với quá trình tái bản và tái tổ hợp DNA chứng tỏ luôn có sự phối hợp chặt chẽ giữa nhiều cơ chế di truyền
III. Sửa chữa và bảo vệ DNA1. Sửa chữa tức thời trong sao chép (cơ chế đọc sửa) Các sai sót trong QT tái bản AND (bắt cặp sai) được nhận biết và sửa chữa nhờ các ADN polymerase Enzyme AND pol có hoạt tính polymerase (tổng hợp) và hoạt tính 3’ exonuclease (phân hủy ADN từ đầu 3’). Trước khi Nu mới được gắn vào DNA pol dò lại cặp base cuối nếu chúng không bắt cặp phản ứng polymer hóa sẽ dừng lại Cặp nucleotide sai cuối đầu 3’ bị loại nhờ enzim DNA polymerase (Exonuclease). Sau khi sự bắt cặp của sợi kép đã đúng polymer hóa được tiếp tục.
2. Các cơ chế sửa sai của tế bào Tóm tắt một số cơ chế sửa chữa DNA như sau: Các phương thức sửa chữa và phục hồi khác nhau loại bỏ sai hỏng
2.1. Quang tái hoạt hóa - Kiểu sửa chữa trực tiếp loại bỏ đơn giản và phục hồi các sai hỏng. - Nhờ các enzim phụ thuộc ánh sáng (light dependent enzime) khôi phục lại các liên kết cộng hóa trị. - Sai hỏng trên DNA do tia tử ngoại gây ra biến dạng cấu trúc xoắn của DNA được sửa và phục hồi nhờ enzim photolyase - Enzim photolyase sử dụng ánh sáng thay đổi liên kết hóa học Nu trở lại bình thường. - Phổ biến ở thực vật, procaryota và eucaryota . Tuy nhiên chưa thấy ở động vật có vú và người.
2.2. Sửa chữa sự ghép đôi lệch giữa các sợi DNA Sửa chữa được tiến hành khi phát hiện bắt cặp sai của baseN • Phương thức sửa chữa: cắt bỏ nhờ một enzim nhận biết base sai hỏng thực sự hoặc thay đổi chiều hướng trong không gian. • Phương thức này phát hiện ở E.coli, nấm men và tế bào động vật có vú.. Trên E.coli có 3 hệ thống enzime khác nhau được sử dụng sửa chữa các sai lệch
2.3. Sửa chữa bằng phương pháp cắt bỏ2 hệ thống a. Hệ thống trực tiếp cắt base sai hỏng thay thế nó trong phân tử DNA. Các bước: • E N glycosinlase nhận biết base bị biến đổi, hay mất gốc amin, hoặc biến dạng cấu trúc xoắn do sai lệch. • Cắt bỏ base ra khỏi phân tử nhờ thủy phân liên kết giữa base – đường. • Enzime DNA polymerase đưa các nucleotid bổ sung vào chỗ trống –>nối đầu 3’-OH của nucleotid trước. • Enzime lygase nối N mới với N sau – nối với đầu 5’ – PO4- . • Chú ý: Mỗi base được một loại N- glycosylase nhận biết riêng + Uraxin - glycosylase + Adenin – glycosylase. + Guanin – glycosylase + Cytozin – glycosylase + Thymin – glycosylase.
Phương thức Cắt bỏ base N
b. Hệ thống sửa chữa cắt bỏ nucleotid - Cắt bỏ một trình tự có base sai hỏng tổng hợp đoạn nucleotid mới. Phổ biến ở hầu hết các sinh vật. • Có sự tham gia của nhiều E để kiểm soát E DNA polymerase tìm ra sai hỏng. • Khi phát hiện được sai hỏng E helicase (endonuclease) sẽ cắt 2 phía của sợi đơn mang lỗi. • E Exonuclease loại bỏ đoạn hỏng ra khỏi DNA. • E DNA polymerase sẽ tổng hợp đoạn DNA mới theo NTBS • E ligase nối đoạn mới được tổng hợp với đoạn cũ. Phân tử DNA được hoàn chỉnh
Sửa chữa Cắt bỏ nucleotid
2.4. Hệ thống sửa chữa sai sót trong tái tổ hợp • Các vị trí sai hỏng trong DNA được phục hồi bằng phương pháp tái tổ hợp. => thu được một bản sao khác của trình tự từ một nguồn không bị sai hỏng. - Một bản sao mới từ nguồn không bị sai hỏng sẽ thành khuôn để tái tổ hợp sợi mới. Bản sao sau đó được dùng để sửa chữa vị trí hỏng trên sợi bị hỏng. - Khi DNA polymerase gặp một số sai lệch trên sợi DNA, có 2 khả năng xảy ra: + Nucleotid được lấp đầy chỗ trống không theo khuôn mẫu do tái tổ hợp vượt qua chỗ sai . Sai sót vẫn tồn tại + Polymerase dừng lại và huy động khoảng 1000 nucleotid , lấp đầy những chỗ trống bị gián đoạn tạo sợi bổ sung từ sợi chị em do tái tổ hợp sai sót ít hơn nhưng vẫn còn. - Trong hai khả năng trên số sai lệch vẫn còn và sẽ được cắt bỏ sau đó bởi sửa sai trực tiếp hay cắt bỏ.
Sửa sai nhờ Tái tổ hợp và tái bản
2.5.Hệ thống sửa sai SOS- sửa sai ngẫu nhiên - Hệ thống SOS: chứa khoảng 30 gen không liên kết bị ức chế bởi protein LexA. - Khi tế bào bị nhiều tác nhân gây đột biến sai hỏng nghiêm trọngSOS sẽ phục hồi tái bản theo cơ chế ngẫu nhiên” error-prone” Ví dụ: phân tử DNA bị sai hỏng nặng nề do chiếu tia tử ngoại hoặc tác nhân ung thư2 sợi DNA đều bị đứt, gẫy thông tin di truyền mất hoàn toàn không có khuôn tái bảnDNA được sửa sai “ngẫu nhiên” duy trì được hoạt động, DNA đột biến. - Hoạt động protein LexA chịu sự kiểm soát của gen recA. + Khi gen recA bị kiểm soát không hoạt động protein LexA hoạt động ức chế hệ thống SOS. + Khi gen recA bị hoạt hóa gây phản ứng phân giải LexALexA bị kích thích thay đổi cấu hình, tự cắt mất hoạt tính ức chế gen của hệ thống SOS được mở.
Cơ chế tác động của protein LexA- recA