Download
1 / 21
210 likes | 220 Views
This article explores the immune response of insects to pathogens, focusing on the recognition of pathogens, communication between immune cells, and the production of antimicrobial compounds. It also discusses the role of symbiotic bacteria in killing infected insects and the interactions between parasites and potential lethal parasites. The article highlights the specificity of entomopathogenic nematodes and the role of parasitoids in the colonization process.

E N D
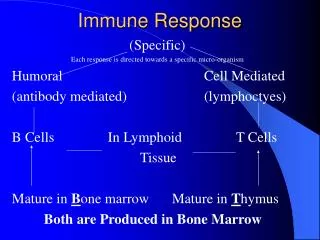
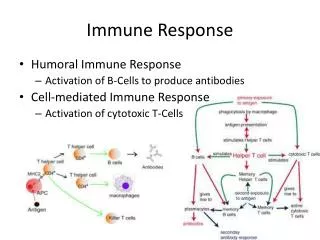
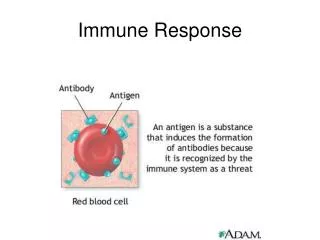
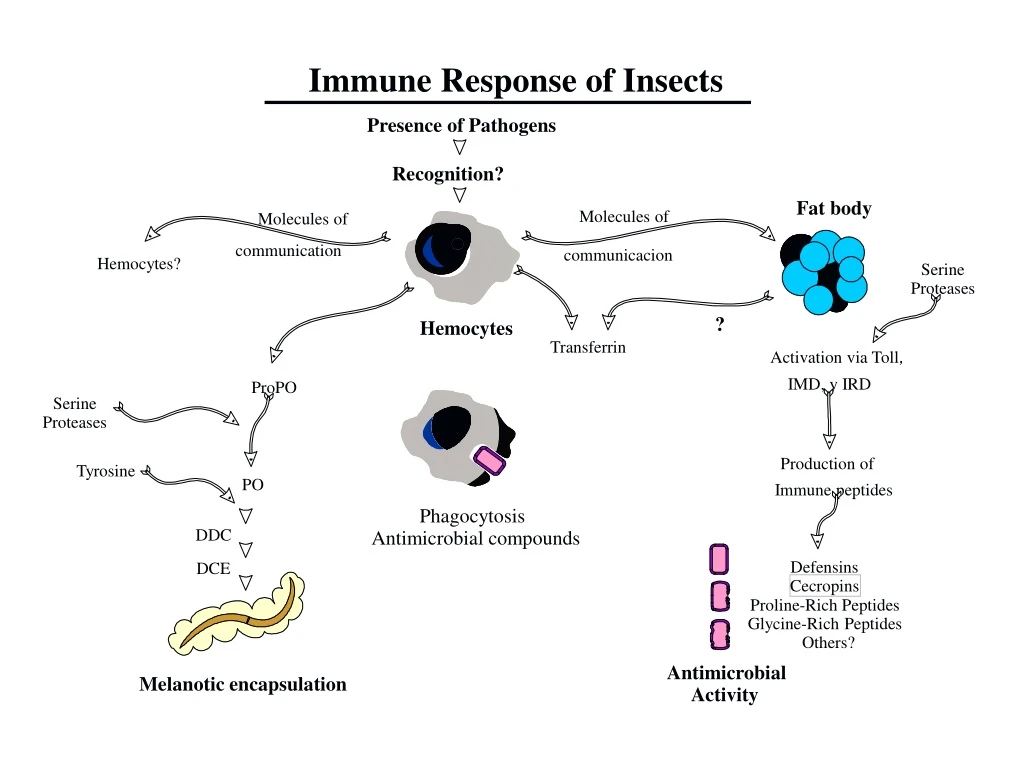
Immune Response of Insects Presence of Pathogens Recognition? Fat body Molecules of Molecules of communicacion communication Hemocytes? Serine Proteases ? Hemocytes Transferrin Activation via Toll, IMD, y IRD ProPO Serine Proteases Production of Tyrosine PO Immune peptides Phagocytosis DDC Antimicrobial compounds Defensins DCE Cecropins Proline-Rich Peptides Glycine-Rich Peptides Others? Antimicrobial Melanotic encapsulation Activity
Rhodnius prolixus Anopheles gambiae T. Cruzi P. falciparum
All of this is based on the concept that the parasite/pathogen can get into a host. In vectors- enter with a bloodmeal from an infected vertebrate. How else can parasites/pathogens get in?
What do you do if you want to get into a host, but when you do you are killed. Who can you enlist to help you? In the colonization process: “the enemy of my enemy is my friend”. Who could you ask for help? 1) others of your same species 2) others from different, but related species? 3) other distantly related species?
What about parasitoids? What about other potential lethal parasites? Why are these invaders not killed by components of the innate immune response?
Lepidopteran larvae-caterpillars Nematode parasites enter but are killed by components of the innate immune response of the insects. Bacteria cannot get into the larvae with any predictability Nematode + bacteria= potent pathogenic duo.
Nematode-Bacterium Complex Entomopathogenic nematodes Nematode-bacterium specificity Lepidopteran host specificity
The entomopathogenic nematodes from the genus Heterorhabditis and Steinernema have entered a symbiotic relationship with bacteria, apparently independent from one another. These bacteria belong to the Enterobacteriaceae, and are closely related to Escherichia coli, our intestinal bacteria, yet they are not harmful to man as they do not grow at temperatures above 35°C. These symbiotic nematodes are able to kill the infected insects fast. Other related species with no symbiotic bacteria penetrate the insect larvae, but they have to wait until the insect dies. Yet other species only attack dead insects. Due to the symbiosis, entomopathogenic nematodes can use the living insect as an energy resource, and the symbiotic bacteria who would not enter body cavity of the insect without the nematodes.
Once in the insect's blood system the nematodes release the bacteria who proliferate fast and kill the insect in approximately 3 days. The bacteria and the insect tissue digested by them forms the food supply for the proliferating nematodes. The proliferation inside an insect can be several 100-thousand fold.
Steinernema sp.-----Xenorhabdus sp. Heterorhabditis sp. ----- Photorhabdus sp.
Fig. 2. Location of symbiotic bacteria in intestines of their respective nematode hosts. (A) Xenorhabdus nematophila cells located in the intestinal vesicle of Steinernema carpocapsae infective juveniles (magnification approximately 1/2 that B). (B) Photorhabdus luminescens cells located in the anteriors and mid-intestine of Heterorhabditis bacteriophora nematodes. In both panels, the images are overlays of epifluorescent and light (Nomarski in B) micrographs. The bacteria are fluorescent due to heterologous expression of the green fluorescent protein (GFP). The bacteria are denoted by arrows and the anteriors of the nematodes are to the top of the figure.
The nematode then feeds on the reproducing bacteria. Commercial products contain the infective juvenile stage of various species. Each species and strain of nematode seems to be most active against a rather narrow groups of insects.
Bio-insecticide products based on entomopathogenic nematodes consist of the infective juveniles stage of the EPN life cycle. Infective juveniles can survive in moist soil for extended periods of time, but do not feed, surviving on stored energy reserves until a host is located. The infective juveniles invade the host, via natural body openings (mouth, anus, spiracles). They penetrate the hemocoel. The infective juveniles release the bacterium into the hemolymph. The bacteria rapidly multiply and the toxins produced kill the insect by septicemia. The nematodes feed upon the bacteria and degraded insect tissue and develop to first generation adult males and females, 2-3 generations. As the insect resource becomes exhausted, most of the juveniles differentiate into third stage juveniles, to become the survival form of the life cycle. The insect cuticle then ruptures and the third stage ensheathed juveniles escape into the surrounding environment.
Caterpillar-nematode-bacterium Inject E. coli into caterpillar- immune response- bacteria killed. Inject M. luteus into caterpillar- immune response, bacteria killed. Inject nematode into caterpillar- nematode killed. Inject associated bacterium into caterpillar- caterpillar killed
The Campoletis sonorensis polydnavirus replicates and is assembled only in the calyx cells of the female oviducts. CsPDV is injected along with the egg and venom at the time of oviposition into the larval host. CsPDV disables the normally vigorous cellular and humoral responses against the egg. After hatching, the endoparasitic wasp larvae develops within the hemocoel of the lepidopteran host, before emerging to spin a cocoon and pupate.
Melanotic encapsulation (A) CsPDV effects upon the defensive melanization pathway. (B) CsPDV effects upon the cell-mediated encapsulation response. Encapsulation includes recruitment of plasmatocytes, capsule formation and granulocyte mediated capsule completion. (C) CsPDV effect upon the induction of antibacterial proteins from the fat body and hemocytes. Hatched bars represent points in the three response pathways inhibited by CsPDV. Abbreviations: NADA, N-acetyl dopamine; DT, dopachrome tautomerase; DDC, DOPA decarboxylase; PO, phenoloxidase; DAT, dopamine acetyl transferase; QMI, quinone methide isomerase; QI, quinone isomerase; ROS, reactive oxygen species; Rel X, rel-like transcription factor.
Polydnaviruses are symbiotic proviruses of some ichneumonid and braconid wasps that modify the physiology, growth and development of host lepidopteran larvae. Polydnavirus infection targets neuroendocrine and immune systems, altering behavior, stunting growth, and immobilizing immune responses to wasp eggs and larvae. Polydnavirus-mediated disruption of cellular and humoral immunity renders parasitized lepidopteran larvae suitable for development of wasp larvae as well as more susceptible to opportunistic infections. Evidence from the Campoletis sonorensis polydnavirus system indicates that the unique genomic organization of polydnaviruses may have evolved to amplify the synthesis of immunosuppressive viral proteins. Immunosuppressive viruses have been essential to elucidating vertebrate immunity. Polydnaviruses have similar potential to clarify insect immune responses and may also provide novel insights into the role of insect immunity in shaping polydnavirus genomes.
The capacity of virulent strains of the endoparasitoid L. boulardi to circumvent melanotic encapsulation is attributed to parasitoid-derived ISFs introduced into host larvae of D. melanogaster. ISFs may target one or more elements of the cellular immune cascade, including melanogenesis, which involves Toll pathway regulation of Spn27A.
Some organisms have formed alliances with unrelated organisms in a mutualistic relationship to allow for a combination that allows a parasite relationship to exist. How did this evolve? Coevolution ???