Download

1 / 35
350 likes | 507 Views
F 1 .F o -ATP synthasa. F 1 .F o -ATPasy „H + -translocating ATPases“ membrána Mtch, baktérií, tylakoidů E 1 .E 2 -ATPasy „Na + /K + - translocating ATPases“ eukaryotické plasmatické membrány A o A 1 -ATPasy archebaktérie – hybridy V a F 1 .F o -ATPas

E N D
F1.Fo-ATPasy „H+ -translocating ATPases“ membrána Mtch, baktérií, tylakoidů E1.E2-ATPasy „Na+/K+ - translocating ATPases“ eukaryotické plasmatické membrány AoA1-ATPasy archebaktérie – hybridy V a F1.Fo-ATPas V-ATPasy protonové pumpy interních membrán (tonoplast, synaptické vesikly neuronů) P-ATPasyATP-depend. pumpy nabitých látek Skupiny ATPas
1937 - Herman Kalckar (DEN) – souvislost ATP synthasy a respirace 1961 - Ephraim Racker (USA) - isolace části F1 ATP synthasy 1961 - Peter Mitchell (UK) – chemiosmotická hypotéza 1964 - Paul D. Boyer – syntéza ATP strukturálními změnami v ATPase 1973 - Boyer – „Binding Change Mechanism„ (energeticky náročným krokem je uvolnění ATP z enzymu 1981 - John E. Walker – sekvence genu pro ATPsynthasu. 1994 - Walker a spol. – struktura F1-podjednotky 1996-1997 – prokázány rotace během syntézy ATP * chemicky (Richard Cross, USA) * spektroskopicky (Wolfgang Junge, Germany) * mikroskopicky (Masasuke Yoshida, Japan). ATP synthasa: Přehled historie
Ohlédnutí za historií ATP synthasy … Boyer Racker Slater Mitchell
ATPasa: Nobelova cena 1997 Paul Boyer – „binding change mechanism“ John Walker – krystalizace enzymu a struktura
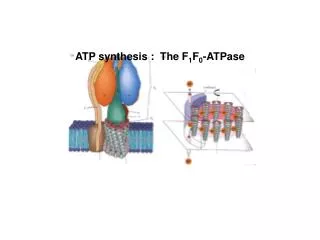
ATP synthasa – E. coli • Fo podjednotka • (160Da) • řetězce a,b,c • F1 podjednotka • (370kDa) • řetězce , , , , Stechiometrie: a.b2.c9-12.3.3...
ATP synthasa Model struktury ATPasy z E.coli
ATP synthasa - MTCH Podobná struktura x souvislost různých podjednotek -řetězec mtch -řetězec E.coli
ATPasy – c-řetězce v elektronovém mikroskopu ATPasa z rostliných thylakoidů
ATPasa – enzymologie DCCD (dicyklohexylkarbodiimid) * vysoce specifická a účinná vazba na c-řetězec * blokace H+ vodivosti Fo * kovalentní vazba s konzervovaným Asp/Glu v transmembránovém helixu c-řetězce * vazba 1 molekuly DCCD blokuje kompletní enzym = kooperace a funkční závislos c-podjednotek
-kovalentní vazba vlákna aktinu s připevněným fluoroforem ATP synthasa – důkaz rotace
ATP synthasa – důkaz rotace * -podjednotky přichyceny ke sklu His-tagem * po přídavku ATP otáčení ve směru hodinových ručiček
ATP synthasa – vliv délky vlákna aktinu - odpor kladený viskozním prostředím buňky je úměrný druhé mocnině délky vlákna
ATP synthasa – kroková rotace při nízké konc. ATP • ATP 20nM • při nízké konc. ATP probíha rotace po jednotlivých krocích • vazba pouze 1 molekuly • chybné kroky zpět = stochastický pohyb ATP 200 nM
ATPasa – rotace s částicí zlata recorded 8000 fps playback 1/257 s • Připojení částice Au (cca 4 x větší než ATPasa): • rotace 8000 rpm • kroky po 120 , rychlost v jednom kroku cca 120000!
ATPasa – model rotace • Kontrola 2 stavy: • vazba ATP • odstup Pi
ATPasa – model Fo 33 Perspektivní pohled
ATPasa – model Fo 33 Pohled z boku
ATPasa – model Fo 33 Průřez podjednotkou
ATPasa – model Fo 33 Pohled svrchu
ATPasa – model Fo 33 Pohled svrchu
ATPasa – detail aktivního místa Vazebné místo pro ATP na -řetězci
ATPasa – detail aktivního místa Vazebné místo pro ATP na -řetězci (Molscript)
Titul Boční pohled na 1 -podjednotku a 1 -podjednotku
ATPasa – uspořádání c-podjednotek 3D zobrazení prstence c-jednotek
„Inhibiční protein“ – blokování hydrolýzy ATP bez přít. p * anoxie = pokles pH matrix = vyšší afinita k enzymu „Záklapka“ – jednosměrnost pohybu -podjednotky u baktérií * přechod mezi 2 velmi rozdílnými konformace jenom díky otáčení -podjednotky Thioredoxin – zamezení hydrolýzy ATP v thylakoidech ve tmě * tma = nečinnost PS1 = není redukovaný feredoxin pro redukci thioredoxinu – tvorba disulfidického můstky a inaktivní formy ATPasy ATPasa – netermodynamická regulace