Download
1 / 39
390 likes | 469 Views

E N D
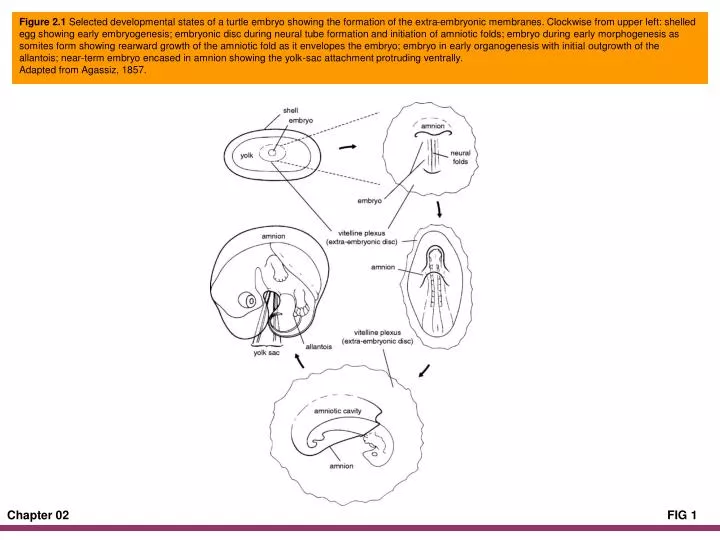
Figure 2.1 Selected developmental states of a turtle embryo showing the formation of the extra-embryonic membranes. Clockwise from upper left: shelled egg showing early embryogenesis; embryonic disc during neural tube formation and initiation of amniotic folds; embryo during early morphogenesis as somites form showing rearward growth of the amniotic fold as it envelopes the embryo; embryo in early organogenesis with initial outgrowth of the allantois; near-term embryo encased in amnion showing the yolk-sac attachment protruding ventrally.Adapted from Agassiz, 1857.
Figure 2.2 Paedogenesis and isogenesis in Ambystoma talpoideum. The life history of A. talpoideum demonstrates the complexities of trait development patterns. The ancestral condition for this species is metamorphosis into a terrestrial salamander in less than one year. Under certain environmental conditions, paedogenesis occurs when metamorphosis is delayed and results in sexual maturation of the individual with retention of larval traits (i.e., the larval morphology) producing paedotypic individuals. Isogenesis occurs when similar early larvae follow different developmental trajectories but ultimately produce similar adults. The adults are termed isotypic individuals. Figure courtesy of S. M. Reilly.
Figure 2.3 The concept of heterochrony can be applied to a wide variety of traits. The New World microhylid, Dermatonotus muelleri, has a tiny head relative to its body and, because other New World microhylids are similar, truncation of head development likely occurred in an ancestor to the clade of New World microhylids. (Luis Gasparini)
Figure 2.4 Body forms of some amphibian larvae arranged by habitat type.
Figure 2.5 Reptiles are tightly coiled inside of eggs prior to hatching. Embryos of Plestiodon fasciatus inside of eggs. Developmental stage 39 (upper); stage 40 (lower). (James R. Stewart)
Figure 2.6 Selected larval stages of a typical anuran. Stage terminology from Gosner (1960).
Figure 2.7 Photograph of the egg of a Geochelone sulcata just beginning to hatch. The arrow points to the emerging egg tooth as it begins to slice through the leathery shell. (Tim Colston)
Figure 2.8 General growth pattern trends for amphibians and reptiles. Top: comparison of indeterminant and determinant growth. When growth is relatively indeterminant, constant growth rate as a juvenile is followed by slower, but continuous growth once sexual maturity is reached. When growth is determinant or asymptotic, a sigmoid pattern during juvenile stages is followed by slower growth after sexual maturity and finally curtailment of growth. Bottom graph: hypothetical growth for an ectotherm in a seasonal environment follows a pattern of rapid growth during equable seasons and greatly reduced or no growth during adverse seasons.
Figure 2.9 Amphibian skin. Cross section through the ventral skin of a marine toad Rhinella [Bufo] marina. Abbreviations: Mg, mucus gland; Pg, poison or granular gland; Sc, stratum compactum; Sg, stratum germinativum; Ss, stratum spongiosum.
Figure 2.10 The tropical toad Rhaebo guttatus has enlarged paratoid glands behind the head as well as many other glands over the body surface. Secretions from the paratoid glands are toxic. (Janalee P. Caldwell)
Figure 2.11 Femoral pores of the male of the lizard Sceloporus undulatus are located along the posterior edge of the underside of the thighs. They appear as lines of black spots. (Laurie Vitt)
Figure 2.12 Diagram of the sequential cellular changes during a single shedding cycle in squamate epidermis. Adapted from Landmann (1986).
Figure 2.13 Anolis punctatus shedding its skin. Note that the old skin separates in several places from the new skin (Laurie Vitt).
Figure 2.14 The arrangement of chromatophores in amphibian skin, called the dermal chromatophore unit. The unit consists of xanthophores, which give yellow, orange, or red coloration; the iridophores, which reflect light and cause bright colors; and the basal melanophores, which have dendritic processes that extend between the xanthophores and the iridophores.
Figure 2.15 Frog skin contains a variety of pigments that often result in bizarre intricate patterns, as in this Amazonian Ceratophrys cornuta (Janalee P. Caldwell).
Figure 2.16 Cranial skeletons of representatives of the three clades of extant amphibians. Dorsal, lateral, and ventral views (left to right) of the caecilian Epicrionops petersi, the salamander Salamandra salamandra, and the frog Gastrotheca walkeri.
Figure 2.17 The hyobranchial skeleton of a typical vertebrate, the salamander Cryptobranchus (dorsal view), and the caecilian Ichythyophis (ventral view).Reproduced, with permission, from Duellman and Trueb, 1986.
Figure 2.18 Cranial skeletons of representatives of the three clades of living reptiles. Dorsal, lateral, and ventral views (left to right) of the turtle Pseudemydura umbrina, the crocodylian Alligator sinensis, and the lizard Ctenosaura pectinata. Adapted from Gaffney (1979), Iordansky (1973), and Oelrich (1956), respectively.
Figure 2.19 Reptile teeth can sit on top of the jaw (acrodont), embedded in the jaw (thecadont), or on the side of the jaw (pleurodont). Tooth location is one of the many important taxonomic characters used to separate major taxa. Adapted from Kardong, 2006.
Figure 2.20 Evolution of skull openings (fenestre) in modern reptiles. Variation exists in the openings (fenestre) behind the orbit and the position of the postorbital (Po) and squamosal (Sq) bones that form the arch from the orbit to the back of the skull. The anapsid (closed) condition is thought to be ancestral. Lizards (including snakes) clearly have modified diapsid (two fenestre) skulls. Turtles, which have been placed historically in the Parareptilia based on the absence of a second fenestra, more likely have a highly modified diapsid skull in which both fenestre have closed. Other bones shown include the quadratojugal and the jugal. Adapted from Kardong, 2006.
Figure 2.21 Evolution of jaw structure and function in squamates. Clockwise from upper left, ancestors of squamates had rigid jaws and skulls such that the skull lifted as a unit when opening the mouth (metakinesis). The "hanging jaw" of squamates (streptostyly) allowed rotation of the lower jaws on the quadrate bone. Scleroglossans have kinetic joints in the skull located behind the eyes (mesokinesis), and snakes have an extra joint located anterior to the eyes (prokinesis). Increased flexibility of the skull allows greater prey-handling ability. The red circle with a cross indicates focal point of rotation.
Figure 2.22 Anterior and lateral views of vertebral morphology of the tetrapods Salamandra salamandra and Crocodylus acutus and a schematic lateral view of an early tetrapod. Adapted in part from Francis (1934), Mook (1921), and Goodrich (1930).
Figure 2.23 Postcranial skeletons (ventral view) of a gray treefrog (Hyla versicolor) and a hellbender (Cryptobranchus alleganiensis). Adapted from Cope (1898).
Figure 2.24 Partial skeleton of a crocodylian showing the variation in structure of vertebrae. The vertebral column is divided into five regions. Note the location of the gastralia (floating "ribs"). Redrawn from Kardong, 2006.
Figure 2.25 Skeleton of a modern turtle showing fusion of vertebrae to the shell. Adapted from Bellairs, 1969.
Figure 2.26 Graphic showing primitive lateral-sequence gait of a salamander. The center of mass (red circle) remains within the triangle of support (dashed line), and three of the four limbs meet the ground at the same time. During a trot gait (not shown), diagonal limbs meet the ground at the same time and the center of gravity falls on a line connecting those limbs. Often, the tail is used to stabilize the trot gait, which forms a triangle of support. Redrawn from Kardong, 2006.
Figure 2.27 Ventral views of the pectoral (upper) and pelvic (lower) girdles of a juvenile tuatara (Sphenodon punctatus). Adapted from Schauinsland, 1903.
Figure 2.28 Above: A diagrammatic lateral view of the brain and spinal cord of a frog. Below: Structure of the frog brain.
Figure 2.29 Infra-red heat-sensing pits are located below and posterior to the nares in pit-vipers. These sense organs detect movement across a thermal landscape based on relative temperature. The snake in the photograph is Bothriopsis bilineata from the Amazon rain forest. (Laurie Vitt)
Figure 2.30 Lateral view of the anatomy of a lizard's ear. The otic capsule consists mainly of the opisthotic and prootic. Adapted from Baird, 1970.
Figure 2.31 Cross section of the anatomy of a snake's eye. Adapted from Underwood, 1970.
Figure 2.32 Lepidosaurians can have gustatory organs (taste buds), nasal olfactory systems (sense of smell), and/or vomeronasal systems (chemosensory using the tongue to transport chemicals). Adapted from Schwenk, 1995.
Figure 2.33 Lateral view of the circulatory system of a frog.
Figure 2.34 Heart anatomy of a turtle and a varanid lizard; diagrammatic ventral views of frontal sections. The arrows indicate only the general pathway of blood flow through the ventricle into the aortic arches. Adapted from Burggren, 1987.
Figure 2.35 A variety of glands occur in the oral region of the head of reptiles, although not all reptiles have all glands shown. Premaxillary, nasal, and palatine glands secrete mucous to lubricate the mouth. Lacrimal and Harderian glands secrete fluids that wet the vomeronasal region and the eyes. The Duvernoy's gland occurs in venomous snakes and produces venom.
Figure 2.36 Visceral anatomy of a generalized male snake; a ventral view.
Figure 2.37 Internal morphology of generalized reptilian lungs; schematic cross sections of a single-chambered lung (top), a transitional lung (middle), and a multichambered lung (lower). The central chamber of a single-chambered lung is not divided by a major septum, although small niches are commonly present along the wall. The transitional lung has a central lumen partially divided by large septum. The multichambered lung is partitioned into numerous chambers of various sizes; all chambers communicate with the intrapulmonary bronchus via an airway. Adapted from Perry, 1983.
Figure 2.38 Ventral view of the reproductive tracts of a female (left side) and male (right side) salamander.
Figure 2.39 Schematic lizard showing the location of some digestive and endocrine glands.