Download
1 / 16
160 likes | 179 Views
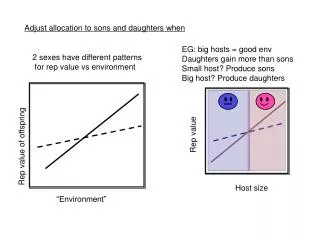
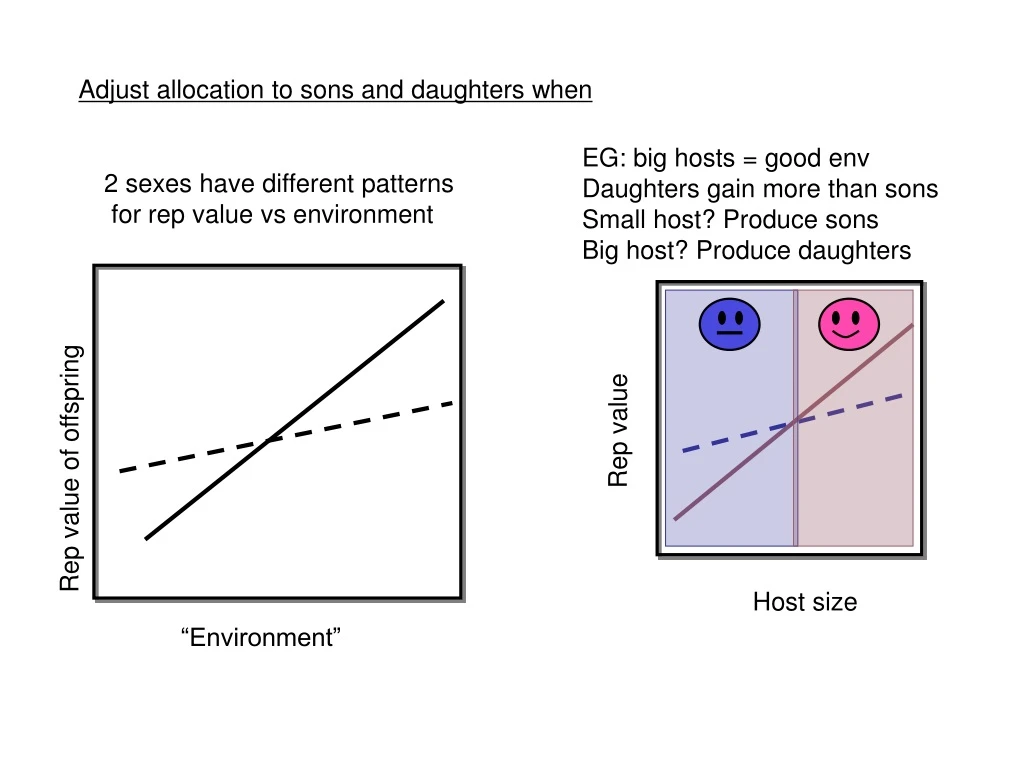
Adjust allocation to sons and daughters when. EG: big hosts = good env Daughters gain more than sons Small host? Produce sons Big host? Produce daughters. 2 sexes have different patterns for rep value vs environment. Rep value. Rep value of offspring. Host size. “Environment”.

E N D
Adjust allocation to sons and daughters when EG: big hosts = good env Daughters gain more than sons Small host? Produce sons Big host? Produce daughters 2 sexes have different patterns for rep value vs environment Rep value Rep value of offspring Host size “Environment”
Concepts • Population (overall) sex ratio [prop. Individuals that are males] • Population (overall) investment (allocation) ratio [prop. Resources invested in males Frank’s F/M different? ] • Complications due to life history details multiple pressures resource distribution current-future trade offs fecundity population density habitat
Population level Offspring SRs at the individual level should be biased but Fisher still holds at the population level If the population SR is 0.5 then individuals are not biasing offspring SRs Very hard to predict the population investment ratio… Can make predictions about relative differences in SR / investment across species / populations Population SR should be biased to the sex produced in the “poorer” conditions / cheaper to make / first sex
1. Population SR should be biased to the sex produced in the “poorer” conditions This is true: #males*Wmales = #females*Wfemales Because all offspring have a mum & dad EG: big hosts = good env Daughters gain more than sons Small host? Produce sons Big host? Produce daughters - Sons cheaper (less food provisioning req’d ) - Expect male biased population SR - Rep value of females increases with investment faster than for males - So, Wmales < Wfemales therefore, #males > #females - Otherwiseeqn doesn’t balance Rep value Host size
2. Population investment (allocation) ratio Relative allocation of overall popn resources to males and females Why is this interesting? - hard to quantify - not meaningful when ESD - hard to make predictions (eg fitness functions * resource distribution) [Not obvious that investment ratio and popn SR can be predicted from each other?] Females produced when lots resources. Say, resource requirement lowers: -> more females than males produced -> males lose largest size class + average size of females decreases Both sexes get smaller, but bigger effect on males[?] so size ratio increases [which drives which? sex ratio?]
3. Interesting things with sex change Popn SR biased to sex that individuals mature as Popn SR less biased if early maturers of second sex are present Popn SR more biased to first sex when that is females
3.1 Interesting things with sex change Popn SR biased to sex that individuals mature as Popn SR less biased if early maturers of second sex are present Popn SR more biased to first sex when that is females i.e. cheaper sex, poorer conditions… same logic Satisfy #males*Wmales = #females*Wfemales
3.2 Interesting things with sex change Popn SR biased to sex that individuals mature as Popn SR less biased if early maturers of second sex are present Popn SR more biased to first sex when that is females Same logic again Eg: sps where females -> males + directly developed males - Direct males have lower rep value than sex change males - Lowers the average rep value of males - So fewer females req’d to balance #males*Wmales = #females*Wfemales • (compared to previous scenario) • Support in fish when females -> males • Not when males -> females. Why? • (expect fewer males when direct females are present) • - males may have higher mortality risk than females [fertility insurance?] • - strange life histories, hard to sample etc
3.3 Interesting things with sex change Popn SR biased to sex that individuals mature as Popn SR less biased if early maturers of second sex are present Popn SR more biased to first sex when that is females (protogynous) Same logic again #males*Wmales = #females*Wfemales Includes differences in size - fitness functions for males Female fecundity increases rapidly with size Female -> male (protgyn) Male rep value has to increase even faster At size of sex change, rep value increases fast Wfemales <<< Wmales Male-> female(protand) Male rep value does not vary so much with size At size of sex change, rep value increases less Wmales <Wfemales Rep value 1 size Rep value 1 size
4. Confusion with environmental Sex determination • Sex determined by individuals response to environmental cue: temp, light, food… • Offspring better able to deal with information than parents • Development in different environments has diff fitness consequences for sexes • either by directly affecting fitness • or by correlating with a scenario/environment, current or future, that does • - T+W applied to ESD is called the ‘differential fitness model’ Gammarus: Sex determined by photoperiod Male if hatch in short day photoperiod, Female if long day. Males develop sooner so have longer to grow before breeding season Fitness gains from large size are bigger for males than females - females have more eggs if large, but more importantly… - large males more likely to get mate and more likely to be a large female
4.1 ESD in bigger and longer lived animals Differential fitness seems to explain Gammarus but not resolved for much else - especially temperature sex determination, widespread across reptiles • eg leopard geckos: • cool + hot temperatures -> females • males only at intermediate temps • eg sea turtles: • cool temperatures produce males • narrow range for mixed sex clutches • females from warm temps Prop. male Prop. male Incubation temp Incubation temp
4.2 The problem of ESD Can’t explain why ESD evolved in most cases (not clear if differential fitness) Can’t understand / predict sex ratio evolution [possible problem with climate change] • Hard to do experiments to answer questions, eg: • as incubation temp confounded by other things, like sex! • hard to quantify fitness costs / benefits in long lived things • reason for / mechanism of ESD inferred from SR [seems a bit backwards] • Too hard to test or we are missing something?
4.3 Explanations for why environmental Sex determination occurs • Differential fitness (~T+W) • TSD is ancestral, not enough genetic variation or selection for GSD - heritable variation for TSD - taxa with a mix of GSD and TSD - lots of shift between TSD and GSD in phylogeny • Remove risk of inbreeding between sibs • single sex clutches generally not much more frequent than mixed • females lay multiple clutches per season & long lived • single sex clutches do not need TSD to occur • TSD species not more inbred than GSD • LMC - not correct ecology / life histories - TSD species not more inbred - don’t need to evolve TSD to have female biased clutches
4.4 Fitting the differential fitness hypothesis • Incubation temperature has differential fitness consequences • phenotype, survival, longevity • - sex specific temperature optima? • eg male skinks from cool temps run faster and females from warm • temps run faster • - suffer least from bad environment? • eg map turtles from cool temps are a bit rubbish • - benefit most from good environment? • classic T+W Incubation temperatures correlates with something important - signal season and hence time for growth in certain conditions - only likely to be important in short lived animals Incubation temperature doesn’t matter - mothers use it to allocate offspring sex - in response to condition (FW turtles) or popn SR - not compelling
4.5 What about the consistently useful popn SR prediction ? If differential fitness is true, then “popn SR biased to sex from poorer environment” eg sea turtles Populations very female biased (< 5% males) Females produced from warm temperatures - are these bad temperatures or does the theory not hold here? Good case where other complexities are important when thinking about popn SR eg nest site fidelity in sea turtles • Females return to natal beach • - more females produced from warmer beaches, • warmer beaches have more females return to them • thus produce even more females • Selection to produce sons • requires earlier nesting, [climate change issues] • or lower nest site fidelity [risky as natal site obviously successful] • enough genetic variation?
4.6 What should be done? • Mixture of approaches • manipulate temperature independently of sex, eg hormones, GSD species • - comparative approach, esp in species with more sexual dimorphism • New hypothesis • sex ratio + sex determination mechanism + nest site fidelity + genetic conflicts? • [maybe they all have a temperature activated SR distorter!] • Separation of evolution vs maintenance [possible multiple explanations] • [Learn something about sex determination, other temp sensitive genes?]