Download
1 / 55
680 likes | 1.02k Views
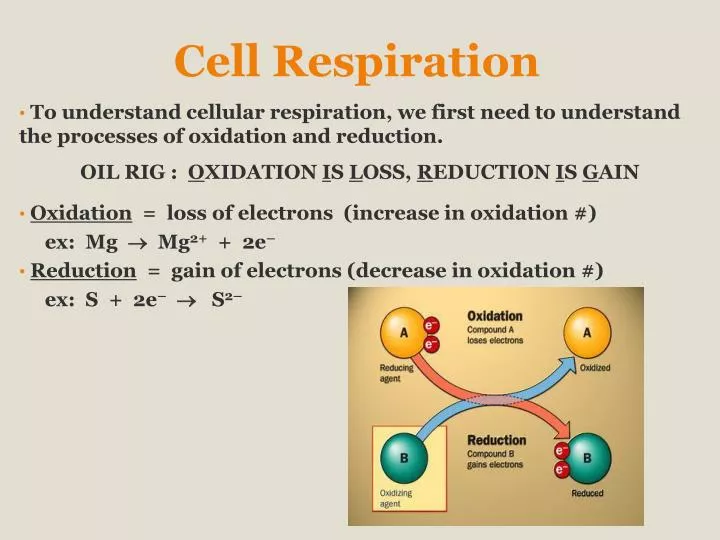
Cell Respiration. To understand cellular respiration, we first need to understand the processes of oxidation and reduction. OiL RIG : O xidation I s L oss, R eduction I s G ain Oxidation = loss of electrons (increase in oxidation #) ex: Mg Mg 2+ + 2e –

E N D
Cell Respiration To understand cellular respiration, we first need to understand the processes of oxidation and reduction. OiL RIG : Oxidation Is Loss, Reduction Is Gain Oxidation = loss of electrons (increase in oxidation #) ex: Mg Mg2+ + 2e– Reduction = gain of electrons (decrease in oxidation #) ex: S + 2e– S2–
Oxidation & Reduction In biology: oxidation often involves gaining oxygen or losing hydrogen reduction often involves losing oxygen or gaining hydrogen
Oxidation & Reduction Examples of biological oxidation-reduction reactions:
Cell Respiration Cell respiration is the controlled release of energy from organic compounds in cells in the form of ATP. Chemical bonds store energy. When we burn things (like wood or gasoline) we release that stored energy, in a process called rapid oxidation. This process is not controlled by enzymes, and results in the breaking of many, many covalent bonds in a very short period of time, and thus a nearly uncontrolled release of energy.
Cell Respiration Cells break down (metabolize) their organic nutrients by slow oxidation, through a series of enzymes. These function of these enzymes is to catalyze a sequence of reactions in which the covalent bonds are broken (oxidized) one at a time. Each time a covalent bond is broken, a small amount of energy is released. The ultimate goal of releasing energy in a controlled way is to trap the released energy in the form of ATP molecules.
Cell Respiration There are two types of cellular respiration: aerobic cell respiration – breaks down glucose in the presence of oxygen to give a high yield of ATP (34-38 ATP) anaerobic cell respiration – breaks down glucose in the absence of ATP to give a low yield of ATP (2 ATP) Let’s look at a quick overview of aerobic cellular respiration…
Aerobic Cell Respiration In aerobic cellular respiration: Glucose is oxidized (loses protons/hydrogen & electrons to oxygen). Oxygen is reduced (gains protons/hydrogen & electrons from glucose).
Assuming that glucose is the organic nutrient being metabolized, both types of respiration begin with the same process: glycolysis. (Keep in mind, though, that respiration can also begin with amino acids or fatty acids.)
Glycolysis In glycolysis, glucose (a 6-carbon sugar, or hexose) in the cytoplasm is broken down into two molecules of pyruvate (a 3-carbon sugar, or triose). Some, but not all, of the covalent bonds in glucose were broken during this process, and the energy from these bonds was used to form a small number of ATP molecules. Two ATP molecules were needed to start the process of glycolysis (the energy investment phase), and four ATPs were produced (the energy payoff phase), for a net yield of two ATPs during glycolysis. Glycolysis occurs in the CYTOPLASM. http://leavingbio.net/respiration-(higher%20level).htm
Looking at this process in detail, there are 4 phases of glycolysis: 1. PHOSPHORYLATION Phosphate groups from 2 ATPs are used to phosphorylate glucose into fructose-1,6-bisphosphate 2. LYSIS The fructose is split into two 3-carbon sugars called glyceraldehyde-3-phosphate (G3P) 3. OXIDATION Each G3P is oxidized by giving up a proton (H+) and 2 electrons (e-) to NAD+, reducing NAD+ to NADH. As NADH is formed, energy is released. This energy is used to add an inorganic phosphate group (Pi) to the 3-carbon compound, so that each now has 2 phosphate groups. 4. ATP FORMATION Enzymes remove all 4 phosphate groups (2 from each 3-carbon compound) and add them to 4 ADP molecules to form 4 ATP molecules. The 3-carbon molecule that remains is called pyruvate or pyruvic acid.
Non-protein components of enzymes that are required for the enzyme to function are called cofactors. If the cofactor is an organic compound, it is called a coenzyme. Coenzymes make up a part of the active site, which is why the enzyme won’t work without them. NAD+stands for nicotinamide adenine dinucleotide. NAD+ is formed from niacin (vitamin B3). Among its other roles, NAD+ is needed in its reduced form (NADH) for the first enzyme in the electron transport chain. NAD + H+ + 2e- NADH Animation of how NAD+ works: http://highered.mcgraw-hill.com/sites/0072507470/student_view0/chapter25/animation__how_the_nad__works.html
This metabolic pathway (glycolysis) is controlled by enzymes. When ATP levels are high in the cell, feedback inhibition will block the first enzyme of the pathway (formation of PFK). This inhibition will slow or stop the process of glycolysis. Then, as the ATP supply starts running low in the cell, the enzymes become unblocked (ATP is removed from the enzyme because it is needed by the cell), thus the enzymes are no longer inhibited, and the glycolysis pathway proceeds, allowing more ATP to be made.
Glycolysis – Summary 2 ATPs are needed to start the process of glycolysis. 4 ATPs are produced; a net gain of 2 ATPs. 2 molecules of NADH are produced. Glycolysis involves 4 major processes: substrate-level phosphorylation lysis oxidation ATP formation Glycolysis occurs in the cytoplasm. Glycolysis is an enzyme-controlled metabolic pathway that is regulated by feedback inhibition. The main contribution of glycolysis to cell respiration is the production of two pyruvate molecules, which will be used in the next phase of cell respiration.
Types of Phosphorylation We should probably differentiate between the two types of phosphorylation you need to know: Substrate level phosphorylation occurs when a phosphate group is taken directly from a substrate and transferred to ADP to make ATP. Oxidative phosphorylation occurs when electrons are transferred from the oxidation of NADH and FADH2 to molecules of O2. These high-energy electrons were used to create a proton (H+) gradient, and the energy from this gradient was harnessed to make ATP from ADP + Pi. (In other words, the inorganic phosphate was just floating around free, and not removed from some substrate molecule.)
Glycolysis – Summary At the end of glycolysis, you have two molecules of pyruvate (pyruvic acid). This pyruvate has one of two possible metabolic fates that we will consider. If oxygen is present in the cell, the pyruvate can enter the mitochondria and continue on with aerobic respiration. If no oxygen is present in the cell, the pyruvate remains in the cytoplasm where it undergoes anaerobic respiration (also called fermentation). Let’s consider anaerobic respiration first, of which there are two types: lactic acid fermentation & alcoholic fermentation.
Alcoholic Fermentation All organisms begin cell respiration with glycolysis. At the end of glycolysis, there are two molecules of pyruvate, and 2 ATPs have been produced for energy for the cell. Yeast is a common once-celled fungus that uses alcoholic fermentation to make ATP. Yeast is a facultative anaerobe (it only performs anaerobic respiration when no oxygen is available; if oxygen is available, yeast will perform aerobic respiration). Some types of bacteria are what we call obligate anaerobes – they can only perform respiration in the absence of oxygen, and are generally poisoned by oxygen.
Alcoholic Fermentation Yeast cells convert both molecules of pyruvate (3C) into molecules of ethanol (2C), plus a molecules of CO2: C6H12O6 2 C3H4O3 2 C2H5OH + 2 CO2 + 2ATP (+ 2NADH + 2H+) In bread production, this CO2 is what causes the dough to rise, and the alcohol evaporates during baking. Note that yeast also recycles the NADH produced during glycolysis back into NAD+ (as you see clearly on the next slide).
Lactic Acid Fermentation When you are performing strenuous labor or exercise, your muscle cells quickly exhaust their stored ATP from earlier aerobic respiration. For example, a sprinter running a 200 m dash uses up all their available ATP supply in the muscles in only 5 seconds. It takes at least 30 seconds after the start of the race before enough extra oxygen breathed in has reached the muscle cells and enough oxidative decarboxylation (CO2 removal) and oxidative phosphorylation (adding Pi to ADP) can occur to replace the ATP. In the interim, your muscle cells would be burning glucose anaerobically in a process known as lactic acid fermentation. (For the first 15 minutes of prolonged activity, your body uses energy stored in glycogen for aerobic/anaerobic respiration. After that, the body begins to burn energy stored as fat.)
Lactic Acid Fermentation Keep in mind that different cells will be undergoing aerobic and anaerobic respiration simultaneously during prolonged activity. The equation for lactic acid respiration in your cells is: C6H12O6 2 C3H4O3 2 C3H6O3 + 2 ATP (+ 2NADH + 2H+) As with alcoholic fermentation, lactic acid fermentation occurs only in the cell’s cytoplasm.
Lactic Acid Fermentation During anaerobic respiration, your muscle cells convert pyruvate into lactic acid. This lactic acid dissociates into lactate and H+ ions: Muscle cells can continue this type of anaerobic respiration at high rates for one to three minutes, during which time lactate can accumulate to high levels. A side effect of high lactate levels is an increase in the acidity of the muscle cells. The same metabolic pathways that break down glucose for energy perform poorly in this acidic environment, and cramps are a natural defense mechanism for the body to prevent permanent damage during extreme exertion by slowing the key systems needed to maintain muscle contraction.
Lactic Acid Fermentation Once the body slows down, oxygen becomes available and lactate reverts back to pyruvate, allowing continued aerobic metabolism and energy for the body’s recovery from the strenuous event.
Anaerobic Cell Respiration Note that in both types of anaerobic respiration (lactic acid fermentation and alcohol fermentation), no additional ATP was produced. Thus, the only ATP yield from anaerobic respiration is the 2 ATP molecules from glycolysis. That’s not much, but some ATP is better than none…
Aerobic Cell Respiration So, aerobic cell respiration is much more efficient than anaerobic respiration, because you get far more ATPs from it (~36 ATP vs. 2 ATP). Without oxygen, the cell’s metabolic pathways cannot fully oxidize the glucose molecule. The ethanol and lactic acid produced during anaerobic fermentation represent unoxidized portions of the glucose molecule. Aerobic respiration leaves no such byproducts. In order for glucose to be fully oxidized, the pyruvate molecules formed during glycolysis must be shipped from the cytoplasm (where glycolysis occurred) into the mitochondrion. This is where the final two stages of aerobic cellular respiration (the Krebs cycle and the electron transport chain) occur.
The Link Reaction Pyruvate enters the matrix of the mitochondrion via active transport. Inside, the 3-carbon pyruvate is decarboxylated (loses a CO2) to form a 2-carbon acetyl group. This CO2 that gets removed diffuses into the blood and we exhale it. The acetyl group is oxidized by NAD+, which removes a proton and two electrons to form NADH (NAD+ is reduced to NADH + H+). Then the acetyl group combines with reduced coenzyme A to form acetyl CoA. This acetyl CoA starting point for the next phase of aerobic cell respiration, called the Krebs cycle.
The Link Reaction When ATP levels in the cell are high, acetyl CoA can be turned into a lipid and stored for later use. When ATP levels in the cell are low, acetyl CoA proceeds on to the Krebs cycle, which occurs in the mitochondrial matrix.
The Krebs Cycle The Krebs cycle begins when the 2-C acetyl CoA combines with a 4-C oxaloacetate to form the 6-C citric acid. (The Krebs cycle is also called the citric acid cycle because citric acid, or citrate, is the first product of the cycle.) You don’t have to know the names of every molecule in the cycle, but you at least need a working idea of how many steps there are in the cycle (8 or 9, depending on how you count), and how many carbons are in the compounds at each step (see next slide). The 6-C citrate is oxidized to a 5-C compound, with the missing carbon released as CO2 from the cell. When the 6-C citrate is oxidized, NAD+ is reduced to NADH + H+. The 5-C compound is then oxidized and decarboxylated itself to form a 4-C compound. Again, a CO2 molecules is released, and NAD+ is reduced to NADH + H+.
The Krebs Cycle As you can see in the previous slide, the 4-C compound undergoes several conformational changes, but remains at 4 carbons, until oxaloacetate is regenerated, and the process starts all over again with a new acetyl CoA. Several other products are produced along the way, though. Another NAD+ is reduced to NADH + H+, and the coenzyme FAD (flavin adenine dinucleotide, shown below) is reduced to form FADH2. Also, one ADP molecule is phosphorylated to form ATP.
The Krebs Cycle It is critical to remember that the Krebs cycle runs twice for every molecule of glucose that enters cellular respiration. Each glucose forms 2 pyruvate molecules, which produce 2 acetyl CoA molecules, and thus two turns of the cycle. So, as a result of the Krebs cycle, for each molecule of glucose, you should produce: 2 ATP 6 NADH + H+ 2 FADH2 4 CO2 You only get 2 ATP from this process, so the main contribution of the Krebs cycle to cell respiration is the production of all those hydrogen carriers/electron carriers (the NADH + H+ and FADH2). These molecules will be crucial to the next phase of respiration, called the electron transport chain.
The Krebs Cycle So far, we only have a net gain of 4 ATPs. Four were made in glycolysis, but two were used to start that process, so glycolysis only netted 2 ATPs. And the Krebs cycle only produces 2 ATPs as well. So the main purposes of the first two stages of aerobic cell respiration are to produce products (pyruvate in glycolysis & electron/proton carriers in the Krebs cycle) that will ultimately be used in the electron transport chain to make the bulk of the ATP. All of these ATPs in glycolysis and the Krebs cycle were produced by substrate-level phosphorylation (transfer of a phosphate group on a substrate directly to ADP to form ATP). The ATP produced in the electron transport chain are made via oxidative phosphorylation, as you will soon see…
ETC & Chemiosmosis The electron transport chain is the only phase of aerobic respiration where oxygen is actually needed. The Krebs cycle occurs in the mitochondrial matrix, whereas the ETC occurs on the mitochondrial cristae (the infoldings of the inner mitochondrial membrane). Embedded on this inner membrane are electron carriers, which are easily oxidized and reduced. The electron carriers are close together and pass the electrons along due to an energy gradient (they have slightly different electronegativities).
ETC & Chemiosmosis Most electron carriers are proteins with iron-containing haem (heme) prosthetic groups, called cytochromes. The haem group of the carriers is the part of the protein that is easily reduced and oxidized. One carrier – coenzyme Q – is not a protein.
In the ETC, electrons pass one carrier to another because the receiving molecule has a higher electronegativity. The sources of electrons moving through the ETC are the coenzymes NADH and FADH2 which were manufactured during glycolysis and the Krebs cycle. Note that FADH2 enters the ETC at a lower PE than NADH, so FADH2 allows production of 2 ATPs, while NADH produces 3 ATPs.
In the process of electron transport, small amounts energy are released. This energy is used to pump protons (H+ ions) from NADH & FADH2 from the matrix out to the intermembrane space.
ETC & Chemiosmosis Since there are so many electron carrier proteins in the ETC, there is only a small difference in electronegativity between each one. Thus, only a small amount of energy is lost at each exchange. This allows the cell to efficiently harness these lower amounts of energy in pumping protons across the inner membrane into the intermembrane space. Protons begin to build up in large quantities between the membranes.
ETC & Chemiosmosis This build-up of protons is part of a process called chemiosmosis. The concentration gradient of H+ ions created by the ETC is harnessed by an enzyme called ATP synthase. Protons are allowed to passively diffuse back into the matrix through this enzyme. The potential energy provided by the movement of these protons down the concentration gradient is harnessed by ATP synthase to phosphorylate ADP into ATP. http://vcell.ndsu.nodak.edu/animations/etc/movie-flash.htm
ETC & Chemiosmosis At the end of the ETC, the electrons have been depleted of energy. The final electron receptor in the ETC is oxygen (O2), because it has such a high electronegativity. This O2 is the oxygen that you inhale when you breathe. A pair of low-energy electrons from the ETC combine with oxygen and two H+ ions to form water. (Remember that protons have been diffusing back into the matrix through ATP synthase, so there are plenty of H+ ions available for this process.) ½O2 + 2H+ + 2e- H2O About 12% of our daily water needs are met by this type of metabolically-produced water.
ETC & Phosphorylation Consider how ATP formation as part of the ETC and chemiosmosis (oxidative phosphorylation) is different from ATP formation in glycolysis & the Krebs cycle (substrate-level phosphorylation). Substrate-level phosphorylation occurs when a phosphate group is taken directly from a substrate and transferred to ADP to make ATP. Oxidative phosphorylation occurs when electrons are transferred from the oxidation of NADH and FADH2 to molecules of O2. These high-energy electrons were used to create a proton (H+) gradient, and the energy from this gradient was harnessed to make ATP from ADP + Pi. (In other words, the inorganic phosphate was just floating around free, and not removed from some substrate molecule.)
Mitochondrial Structure We also need to consider how the structure of the mitochondria is closely related to its function in the Krebs cycle & ETC. The fluid-filled matrix contains the enzymes for the Krebs cycle. The cristae provide a large surface area for the ETC to function (lots of space for lots of enzymes). The membranes provide a barrier, allowing for proton accumulation between them. The enzymes and other molecules necessary for the ETC are embedded in the mitochondrial inner membrane.