Download

1 / 31
330 likes | 593 Views
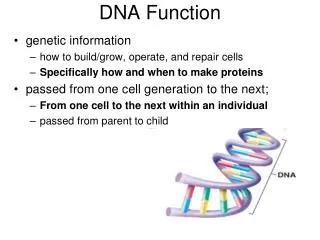
How Histone Post-Translational Modifications Function When They Are Buried Under DNA. Michael Guy Poirier Assistant Professor Department of Physics The Ohio State University. DNA is Highly Wrapped and Compacted in Eukaryotes. Interphase. Mitosis. Luger et al , Nature 1997.

E N D
How Histone Post-Translational Modifications Function When They Are Buried Under DNA Michael Guy Poirier Assistant Professor Department of Physics The Ohio State University
DNA is Highly Wrapped and Compacted in Eukaryotes Interphase Mitosis Luger et al, Nature 1997 Alberts et al, 2002
How can Wrapped DNA be Biologically Active Preinitiation Complex Nucleosome MSH2-MSH6 Richmond and Davey, 2002 Warren et al, 2007 Bushnell, et al Science 2004
Nucleosomes must be altered for DNA Repair and RNA Transcription • Thermal fluctuations (i.e. site exposure) • Chromatin remodeling (i.e. SWI/SNF) • Histone Variants (i.e. CENP-A) • Chromatin associated proteins (i.e. HP1) • Post-translational modifications
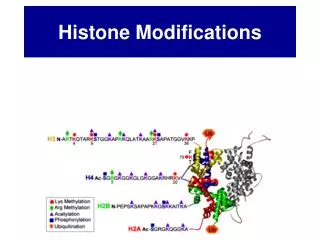
Histone Post-Translational Modifications Histone tails vs. Histone fold domains From Millipore’s website
Modifications in the Histone Fold Domains Key paper: Zhang, Eugeni, Parthun and Freitas, 2003 Cosgrove et al, 2004
Histone H3 Modifications near the Nucleosome Dyad Acetylation of H3-K115 and H3-K122. Mimicking Acetylation with K to Q mutation.
Biological Relevance of H3-K115Ac and H3-K122Ac • Acetylation of K115 and K122 occur individually and together (Zhang, Eugeni, Parthun and Freitas, 2003). • These residues are important for both transcriptional regulation and DNA repair. Data is from: Hyland et al 2005 & English et al 2006
Hypothesis Lysine acetylation in the nucleosome dyad disrupts DNA-histone interaction. This facilitates nucleosome disassembly, repositioning and/or DNA unwrapping. Test Hypothesis By: Preparing Nucleosomes with Acetylated H3-K115 and H3-K122 Using Biochemical and Biophysical tools to determine how nucleosomes are altered
Intein Preparing Acetylated H3 by Expressed Protein Ligation H3(1-109) H3(1-109) Cleavage Ligation H3 - Intein Intein H3 H3 thioester H3 peptide
Purified Histone Octamer Purified H3 Dual Mod Dual Mut K115-Ac K115-Ac K122-Ac K122-Ac WT H3 H3 H2A/H2B H3 thioester H4 Unmodified K115-Ac and K122-Ac 11237 11239 13491 13488 13948 13949 15273 15362 12000 14000 12000 14000 6000 6000 16000 16000 8000 8000 10000 10000 m/Z m/Z
Regular Nucleosome Reconstitute with Acetylated H3-K115 and H3-K122 Cy5 mp2-192 Cy5 Nucleosome positioning Sequence (147 bp) DNA arm (10bp) DNA arm (30bp) mp2-247 Cy3 Cy5 DNA arm (50bp) Nucleosome Positioning Sequence (147 bp) DNA arm (50bp) Nuc Peak DNA Peak H3-K122Ac H3-K122Q. Dual Mod. H3-K115Q H3-K115Ac Unmodified Dual Mut. DNA Unmodified Nucleosomes Fluorescence Intensity Dual Mod. Nucleosomes Nuc DNA Fraction Number
Nucleosome Competitive Reconstitutions • Develop by Jonathan Widom and Coworkers. • Two DNA sequences compete for a limited amount of histone octamer during graduate salt dialysis. • At an intermediate salt concentration, an equilibrium is setup between octamer bound DNA and free DNA. • This equilibrium is frozen in as the salt is fully dialyzed away. • The ratio of the nucleosomes to free DNA is proportional to the equilibrium constant, Keq = [nucleosomes]/[DNA]. • This is compared to a control reconstitution with unmodified histone octamer. • From this a relative free energy of binding is determined: • ΔΔG = -RT[ln(Keq modified) – ln(Keq unmodified)]
Acetylation of K115 and/or K122 Reduces DNA-Histone Binding WT Dual Mod Nuc Kequ DNA K115Ac Δ Δ G = 0.35 ± 0.23 kcal/mol K115Ac & K122Ac Δ Δ G = 0.45 ± 0.25 kcal/mol K122Ac Δ Δ G =0.18 ± 0.26 kcal/mol
ΔΔG Depends on DNA Sequence WT Dual Mod Nuc 5S DNA Kequ WT Dual Mod Nuc 5S DNA Δ Δ G = 0.15 ± 0.05 kcal/mol mp2 DNA Δ Δ G = 0.45 ± 0.25 kcal/mol Mp2 DNA
Measuring Site Exposure with Restriction Enzymes TaqαI Hae III Hind III Hha I Pml I mp2 DNA (192 bp) Hha I Taqα I Hind III Pml I Hae III
DNA site exposure is not altered by the acetylation of K115 and K122 DNA digestion with Taqα I Unmodified K115Ac & K122Ac 0 1 2 4 8 16 24 32 0 1 2 4 8 16 24 32 Uncut Uncut Cut Cut TaqαI Hae III Hind III Hha I Pml I mp2 DNA (147 bp) DNA Fraction Uncut Kequ K115Ac & K122Ac Kequ unmodified Time (min) Position from dyad (bp)
Nucleosomes thermally reposition more rapidly with K115 and K122 acetylated. K115Ac & K122Ac Unmodified Time (min) 0 0.5 1 2.5 5 10 15 20 0 0.5 1 2.5 5 10 15 20 DNA Band Fraction DNA Time (min)
Manipulating Nucleosomes with Magnetic Tweezers Moveable permanent magnet Applies the force with field gradient Force S N Magnetic Bead Flow cell Nucleosomes Biotin- Streptavidin Objective Dichroic mirrors Lamp Laser Lens Dig.-Antidig. CCD Lens Dichroics and Band Pass Filters Glass Surface CCD or APDs
Magnetic Tweezers Determine force from thermal fluctuations L L Measurements are done at fixed force.
Force vs. extension for a single DNA molecule Force (pN) Extension (um)
Nucleosome arrays DNA template AvaI BamHI AvaI Streptavidin Biotin 17 nucleosome positioning sequences (3009 bp) DNA arm (1500 bp) DNA arm (1500 bp) DNA Acrylimide Gel BamHI digestion Native Acrylimide Gel Ava I digestion Native Composite Gel [NPS] [HO] 0 .5 .6 .8 1 1.2 1.5 0 .5 .6 .8 1 1.2 1.5 0 .5 .6 .8 1 1.2 1.5
Unwrapping Nucleosomes by Force Bead Height (microns) Time (sec)
K115 and K122 Acetylation increase fraction of histone octamer force induced disassociation Apply 20 pN of force Wait 8 minutes and count the number of unfolding events Repeat Fraction of Nucleosomes Remaining Relax to zero force Wait 3 minutes Extension Number
What have we learned about K115Ac and K122Ac? • DNA-histone binding is reduced by up to ~0.5 kcal/mol. • Reduction in binding affinity depends on DNA sequence. • K to Q mimic does capture all of the effects of acetylation. • Steric bulk is more important than change in charge. • Do not alter site accessibility. • Facilitate nucleosome repositioning. • Facilitate nucleosome disassociation following DNA unwrapping.
Nucleosomes and DNA Mismatch Construct 5S nucleosome positioning sequence mismatch biotin 167 bp 71 bp Streptavidin - + + + + Sucrose Gradient Purification hMSH2-hMSH6 (nM) 100 50 200 0 0 1 2 1 2 hMSH2-hMSH6 + Nuc + MM DNA + Streptavidin Nucs Nuc + MM DNA + Strept Nuc + MM DNA DNA MM DNA
MSH2-MSH6 drives off nucleosomes MM Nuc DNA + hMSH2/6 MM Nuc DNA + SA MM Nuc DNA Streptavidin 0 10 20 30 40 50 60 Time (min) hMSH2/6 MM Nuc DNA + SA + hMSH2/6 MM DNA + SA + hMSH2/6 nucleosome MM Nuc DNA + SA MM Nuc DNA MM DNA + SA biotin 5S mismatch MM DNA
Acetylation of K115 and K122 Facilitates Nucleosome Removal by MSH2-MSH6 WT MSH2/MSH6 unmodified nucleosomes WT MSH2/MSH6 H3-K115Ac-K122Ac MSH2-K675A/ MSH6-K1140A H3-K115Ac-K122Ac
Summary of MSH2-MSH6 and Nucleosomes • MSH2-MSH6 can drive off the histone octamer from a DNA positioning sequence. • The nucleosome disassociation is ATP dependent • Acetylation of K115 and K122 facilitates nucleosome removal
Acknowledgements Poirier Lab: Alex Mooney Justin North Marek Simon Robin Nakkula Ottesen Lab: Jennifer Ottesen Mridula Manohar Annick Edon Fishel Lab: Richard Fishel Sarah Javaid Mark Parthun and Jonathan Widom Funding