Download

1 / 60
730 likes | 1.13k Views
THE COMPLEMENT SYSTEM. DIL – 6 Oct. 31 st 2013 Mohammed El- Khateeb. The Complement System History Concepts Complement Activation Regulation of the Complement System Biological Function of Complement. COMPLEMENT. Historical Background Nuttall Bacteria in guinea pig blood plasma

E N D
THE COMPLEMENT SYSTEM DIL – 6 Oct. 31st 2013 Mohammed El-Khateeb
The Complement System • History • Concepts • Complement Activation • Regulation of the Complement System • Biological Function of Complement
COMPLEMENT • Historical Background • Nuttall Bacteria in guinea pig blood plasma • Result: bactericidal activity • 1889 Buchner serum borne component designated to Alexin • 1894 Pfeiffer and Issaeff • Vibrio cholera Guinea pig • Survived animal rapidly killed the bacteria • Heating the serum from the immunize animals:destroyed the antibacterial activity in vitro • Heated serum showed immunity • the heat-labile alexin activity : Complement
History of complement • Ehrlich – role of ‘complementing’ antibodies in killing of bacteria. • 1895 – Bordet • Subsequent discovery of components • Current knowledge:- • > 30 proteins in plasma + on cell surfaces • ~ 15% of globulin fraction of proteins
Discovery of Complement 1890s Jules Bordet (Institut Pasteur in Paris) observed: 1. Sheep antiserum to the bacterium Vibrio cholerae caused lysis of the bacteria. 2. Heating the antiserum destroyed its bacteriolytic activity. 3. Addition of fresh normal serum, that contained no Abs against the bacterium and was unable to kill the bacterium by itself, restored the ability to lyse the bacteria by the heated antiserum.
Paul Ehrlich in Berlin carried out similar experiments and named the substance complement,defining it as “the activity of serum that completes the action of Ab.” Bordet won the Nobel Prize in 1919 – Complement-mediated bacteriolysis
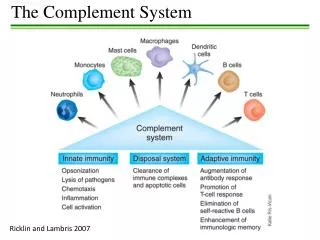
The Complement System Is an enzyme cascade Made Up of numerous Soluble Components • Pro-enzymes, • Receptors • Regulatory Proteins
The Complement System 19 effector components to cascade In tissue fluids & blood & cells) 5 Cell Surface Receptors 9 Regulatory Proteins (Some soluble & some on cell surfaces
Nomenclature • C1 – C1q, C1r, C1s • C4, C2, C3, C5, C6, C7, C8, C9 • Many referred to as ‘zymogens’ • ‘a’ and ‘b’ – added in to denote cleavage products. • ‘b’ – larger fragment • Alternative pathway proteins:- ‘Factors’ or identified by single letters • Complement receptors:- named according to ligand (eg C6 receptor) or using CD system.
The basics! • ‘Innate immune system’ • Cascade • C3 – most important component • Activation:- innate or adaptive systems • Classical:- adaptive immune system – immune complexes bind to C1q • Alternative:- innate – chance binding of C3b to microorganism surface. • Distinction of self from non-self! • Deficiencies:- increased susceptibility to recurrent infections (pyogenic bacteria) OR illnesses a/w production of autoantibodies + immune complexes.
Main roles • Defends against pyogenic bacterial infections • Bridges both the innate and adaptive immunity systems • Assists in disposing of immune complexes etc
Role in Inflammation • Opsonization:- C3b is important! • Chemotaxis:- complement fragments diffuse from target – stimulating cellular movement and activation. • Target cell lysis:-‘membrane attack complex hydrophobic ‘plug’ inserted into lipid membrane bilayer
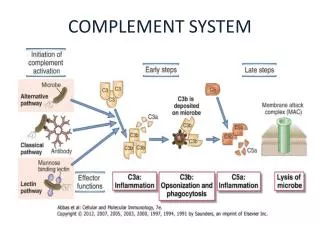
Activation • Pathways:- 1. Classical 2. Lectin 3. Alternative • Common end point: formation of C3 convertase – cleaves to C3a and C3b • Classical + Lectin pathways – C4b2a • Alternative pathway – C3bBb • Ultimately:- converted into C5 convertase - by further addition of C3b. Production of MAC.
Complement Activation 1. Classical Pathway activator IC (immune complex) the main Polymerizer (heparin, polynucleotide) Dextran sulfate Protein (CRP) Liposome Mitochondria of cardiac muscle
Classical Pathway Begins with Ag-Ab Binding Soluble Ag-Ab* or bacteria-Ab* Conformational changes in the Fc portion of Ig Expose a binding site on the CH2 domain of the Fc portion for the C1 complex undergoes conformational change • ‘Autocatalysis’ of C1r • C1s activation * Complement-activating Abs: IgM, IgG1, IgG2, IgG3 (human) C1 molecule C1qr2s2 stabilized byCa++
Difference between C1q binding sites on IgM and IgG subclasses C1q binds to Ag-bound Ab Binding of C1q to Fc induces a conformational change in C1r __ C1r converts to an active serine protease enzyme, C1r__ which cleaves C1s to a similar active enzyme, C1s CH2 domain of the Ab bacterium
C1s hydrolyzes C4 and C2 __ C1s has two substrates, C4 and C2 __ C1s hydrolyzes C4 into C4a and C4b, and hydrolyze C2 into C2b and C2a ___ C4b and C2a form a C4b2a complex, also called C3 convertase, referring to its role in converting the C3 into an active form. an anaphylatoxin*, or a mediator of inflammation C2a C4b ( )
Convertase ____ C4b2a (C3 convertase) hydrolyzes C3 into C3b and C3a __________ C3b binds to C4b2a and formC4b2a3b (C5 convertase) _______ C4b2a3b cleaves C5 into C5b and C5a Opsonization IR IR
Hydrolysis of C3 by C3 Convertase C4b2a formation of a labile internal thioester bound in C3 bind to free - OH or - NH2 groups on a cell membrane generates > 200 C3b C5 Convertase Bound C3b exhibits various biological activities, e.g., binding of C5 and binding to C3b receptors on phagocytes. C4bC2aC3b
The Alternative Pathway Is Ab-independent • The activation of alternative pathway doesn’t need Ab; thus, it is a component of the innate immune system. • It is initiated by cell-surface constituents that are foreign to the host, e.g., bacterial cell wall. • C1, C4 and C2 are not involved in the alternative pathway. • Four serum proteins, C3, factor B, factor D, and properdin, are involved in this pathway.
Complement Activation 2. alternative pathwayactivator LPS bacteria zymosan dextran IgA IgG4 IgE • Does not require Ag-Ab complex formation • Produces active C3 convertase and C5 convertase • Includes four serum proteins • Initiated by foreign cell surface proteins • Active C3 is generated spontaneously
Alternative pathway b. Characteristics: non-specific, rapid distinguish self and non-self C3b positive feedback need a surface to stick or activate C3b
Plasma C3, with an unstable thioester bond, can be hydrolyzed spontaneously into C3a and C3b. • C3b attaches to the surface of bacteria, yeasts, viruses (or even host’s own cells). • Analogous to the C4b2acomplex in the classical pathway Mg++ Ba (stabilization of C3bBb) sialic acid in cellular membrane inactivate C3b
Microbial activation of the Alt C pathway by C3bBb and the control by H and I • C3b in C3bBbbound to host cell or in fluid phase is unprotected therefore the affinity to H factor than B factor so it is susceptible to breakdown by H and I factors. • If bound to bacterial cell the affinity to B is great than bound to H so it will be protected (stabilized) against cleavage
The Lectin Pathway Originates with Host Proteins Binding Microbial Surfaces Lectin: Proteins that bind to a carbohydrate MBL (Mannose-Binding Lectin): - an acute phase protein which binds to mannose residues on glycoproteins or carbohydrates on the surface of microorganisms (structurally similar to C1q) MASP-1 & MASP-2: MBL-Associated SerineProtease (structurally similar to C1r and C1s) Antibody independent activation of classical pathway
MBL is induced during inflammatory responses. • After MBL binds to the surface of a microbe, • MBL-associated serine proteases, MASP-1 and • MASP-2, bind to MBL. • The MBL-MASP-1/2 complex mimics the activity of C1r and C1s, and causes cleavage and activation of C4 and C2. • Thus, the lectin pathway is Ab-independent. It is an important innate defense mechanism comparable to the alternative pathway, but utilizing the elements of the classical pathway, except for the C1 proteins.
C3a increases the inflammatory response by binding to mast cells and causing them to release histamine C5a disperses away from the bacteria. Binds to mast cells and increases inflammation. Most powerful chemotactic factor known for leukocytes
Formation of C5b6789, Membrane-attack Complex C5b attaches to C6, then to C7, and the C5b67 complex inserts into the membrane. binding of C8 to membrane-bound C5b67 induces a 10 Å pore. binding and polymerization of C9, a perforin-like molecule, to C5b678 The completed membrane-attack complex (MAC) has a tubular form and functional pore size of 70 – 100 Å
COPLEMENT ACTIVATION Early steps in activation of classical, lectin, and alternative complement pathways, leading to formation of C3 convertase, C4aC2b in both classical and lectin pathways, and C3bBb in alternative pathway.
COPLEMENT ACTIVATION C3 convertase C5 convertase
COPLEMENT ACTIVATION Formation of MAC. • Late-stage complement components C5b-C9 bind sequentially to form a complex on the cell surface. • Multiple C9 components bind to this complex and polymerize to form poly-C9, creating a channel that disrupts the cell membrane.
Membrane attack complex • Requires enzymatic cleavage of C5 • Sequential binding of C6, C7 (hydrophobic status), C8, C9 (up to 14 monomers) • Formation of lytic ‘plug’ – majority of damage caused by C9 • C9 – analogous to perforin (used by T lymphocytes) • C5b67 – can be inactivated by numerous means (S protein – vitronectinetc) • RBC immunity: poorly lysed by homologous complement • CD59:glycophospholipid foot. Inhibits insertion + unfolding of C9 into membranes.
C5a 70-100 Å C5b The Membrane Attack Complex C5 \ C9 C9 C9 C9 C9 C7 C8 C6
Strict Regulation of the Complement System • Discrimination between microorganisms and self • Passive mechanisms of regulation : highly labile components that undergo spontaneous inactivation if they are not stabilized by reaction with other components • A series of specific regulatory proteins: inactivate various components
Regulation of the Complement System 1. Short half-life 2. Regulation protein Up-regulation: Properdin, C3Nef Down-regulation:C1INH, C4bBp, Hf, If, DAF, CR1, MCP
Regulation of complement pathways • Non-specific attack on “innocent bystander” self- cells • General mechanism: generation of labile components that require stabilization; spontaneous hydroylsis of C3 convertase • Specific mechanisms regulate steps before convertase activity, after C3 convertase activation, and during MAC formation • Ex. C1 inhibitor restricts activation of C4 and C2, RCA proteins (C4bBP, CR1, MCP) • CR1, MCP, and Factor H function in the alternative pathway to bind C3b and prevent C3b association with Factor B to form C3bBb • RCA proteins Cr1, Factor H and DAF (CD55) dissociate C3 convertase • S protein, HRF and MIRL interrupt MAC formation
Downstream effects • C1 – cleaves C4 – forming activated C4b • Two isotypes exist • C4A – binding amine groups (usually on proteins) • C4B – hydroxyl groups on CHO • C4b – allows binding of C2. Acted on by C1s to release C2b. • C4b + C2a = classical pathway convertase (C3) • By definition:- C3 convertase – breaks up C3 to C3a and C3b (focus of further complement activation)
What about regulation? • C1 inhibitor – serine proteinase inhibitor (aka serprin) – binds and inactivates C1r and C1s • Inhibition of formation of C3 convertase enzyme- C4b2a (by ongoing catabolization of C4b by Factor I and C4 binding protein) • Other complement control factors – inhibit complement binding to host cell surfaces • DAF: (Decay accelerating factor) – CD55 • CR1 • MCP: Membrane co-factor protein • Inhibit binding of C2 to C4b; promote ‘decay acceleration’ of C2a from C4b. Assist in catabolism of C4b by Factor I
Regulation… it’s always about rules!!! • Factor H and I • DAF + CR1 – accelerate dissociation of C3bBb ‘How C3b reacts is governed by the surface to which it attaches’ – protected vs non-protected
FIGURE 13.5. Regulators of C3 convertases in (A) classical pathway and (B) alternative pathway.Regulators may dissociate the convertase, cleave the complement component remaining on the cellsurface, or act as a cofactor for this cleavage. C4 binding protein exclusively regulates the classical pathway and factor H regulates the alternative pathway. Factor I, DAF, CR1, and MCP regulate both pathways.
Clinical implications • Complement deficiencies • Glomerulonephritis • C1 inhibitor deficiency • SLE • Paroxysmal Nocturnal Hemoglobiuria PNH • Sepsis • AntiphospholipidAb SyndromeAPLS
Complement deficiency:-Increased susceptibility to pyogenic infections • Contributing factors • Deficient opsonisation • Deficiency compromising lytic activity • Deficient manose-binding lectin pathway • Pyogenic infection:- • Site of defect:- antibody production, complement proteins of classical pathway, phagocyte function • Usually bacteria is opsonised with Ab – complement is then activated, phagocytosis occurs and intracellular killing • Key player:- C3b • Impaired lysis • MAC component deficiency – a/wNeisserial disease* • Risk of meningococcal disease ~ 0.5% / yr (RR 5000 cf normal population) • Deficient lectin • Deficiency occurs due to 1 of 3 point mutations – a/w reduced levels. • Associated with higher risk of infection in children – whilst losing passive immunity • ? Protective against mycobacterial infections
C1 INHIBITOR DEFICIENCY • Autosomal dominant – inadequate production of physiologically adequate C1 inhibitor • Type 1:- 85% - Reduced levels of C1 inhibitor • Type 2:- altered activity • Autoantibodies against C1 inhibitor • Inhibits – C1r and C1s, activated FXI and XII • Consumed by plasmin – trigger for angioedema attacks. • Rx: C1 inhibitor infusion.
Who to Screen? • Hemoglobinuria • Coombs negative haemolyticanaemia • Those with AA or MDS (annual screen) • Haemolyticanaemia • VT without explanation (including unusual sites – eg mesenteric, portal, cerebral etc) • Unexplained arterial thrombosis • Episodic dysphagia or abdominal pain
Functions of complements A. Host Defense B. Disposal of Waste C. Regulation of the Immune Response
Major functions of complement 1 • Production of opsonins 2 2. Production of anaphylatoxins 3 3. Pathogen lysis.
4 4. Enhancement of B-cell responses, 5. Removal of immune complexes, 6. Removal of necrotic cells and subcellular membranes, and responses to viruses. 5 6