Download

1 / 24
240 likes | 542 Views
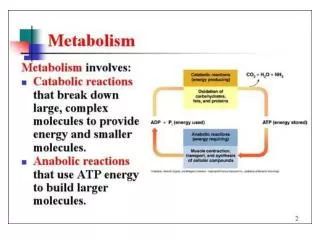
Rasmus Ruus Stom II. Süsivesikute metabolism. Metabolismi põhifunktsioonid inimkehas- Energia omastamine väliskeskkonnast toitainete vormis Toitainete omastamine, lõhustamine (katabolism) ja kasutamine kehaomaste biomolekulide sünteesiks (anabolism).

E N D
Rasmus Ruus Stom II Süsivesikute metabolism
Metabolismi põhifunktsioonid inimkehas- • Energia omastamine väliskeskkonnast toitainete vormis • Toitainete omastamine, lõhustamine (katabolism) ja kasutamine kehaomaste biomolekulide sünteesiks (anabolism). • Senestsentsete biomolekulide lammutamine. • Lõpp-produktide väljutamine. • Organismi sattuvate ksenobiootikumine detoksikatsioon ja väljutamine.
Mis on metaboolne rada ja millised on tema regulatsioonivariandid? • Metaboolne rada on reaktsioonide jada, kus ensüümide toimel muunduvad/tekivad metaboliidid. • Metabolismi regulatsioon on kiire, paindlik ja efektiivne, mis tagatakse rakkudevahelise ja rakusisese kommunikatsiooniga .Põhiprintsiip on teadud valkude/ensüümide aktivatsioon ja inhibitsioon nende fosforüülimise/defosforüülimisega Regulatsioonivariandid on : • Ioonide rakku tuleku kiirus ja raku kompartmentide vahelise liikumise regulatsioon ioonpumpade, kanalite, gradientide abil. • Ensüümide lokalisatsioon koes, rakus • Ensüümide sünteesi induktsioon ja repressioon, sünteesi ja degradatsiooni kiiruse vahekord • Ensüümide kineetiline regulatsioon (aktiv, inhib, modifitseerimine...) • Võtmeensüümide allosteeriline regulatsioon – pos ja neg tagasiside, rist-regulatsioon • Igal metaboolsel rajal on päästikregulatsioon (pöördumatu)
Katabolismi ja anabolismi staadiumid • Aeroobse katabolismi staadiumid: • Makrotoitainete ja senestsentsete biomolekulide lõhustamine monomeerideks, ehitusüksusteks. • Monomeeride, ehitusüksuste muudnamine vähesteks ja lihtsateks metabolismi võtmeühenditeks (monoos > püruvaat > atsetüül-CoA) • Atsetüül-CoA ja Krebsi tsükli komponentide oksüdatiivne lõhustamine lihtsateks lõpp-produktideks. • Anabolismi staadiumid: • Lihtsatest eelühenditest sünteesitakse ehitusüksused/monomeerid • Suuremate biomolekulide ja biomakromolekulide süntees.
Mis on metaboolne energia? Miks ATP on universaalse metaboolse energia vorm? • Metaboolne energia saadakse lõhustatava substraadi energia konverteerumisest ja kasutatakse anabolismis, füsioloogilisteks funktsioonideks, talitluses. • ATP on makroergiliste ühendite põhiesindaja. Makroergilised on ühendid, mille sideme hüdrolüüsi vaba energia standardmuut on üle -25 kJ7mol. ATP-l on see -30,5 kJ/mol. Ta pole kõige makroergilisem fosfaat, aga ta paigutub „supermakroergiliste” ja mittemakroergiliste ühendite vahele
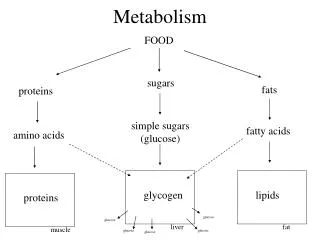
Süsivesikute seedimise ja imendumise põhi-aspektid • Seedimine suuõõnes: algab tärklise hüdrolüüs sülje α-amülaasi toimel. Ensüüm lõhustab sisemisi α(1,4)glükosiidsidemeid • Seedimine maos: amülaas inaktiveerib mao soolhappe ja pepsiini toimel ja maos pole SV seedivaid ensüüme. Niiet seal seda ei toimu. • Seedimine peensooles: mis on ka põhikoht. Pankrease α-amülaas jätkabtärklise seedimist, see toimub duodenumi valendikus. Amülaaside tekitatud produktid lõhustatakse monoosideks. Lõplik hüdrolüüs toimub hariäärise pinnal • SV seedimishäired: maldigestiooni tingib pankrease puudulikkus, mao tugev happesus, ka hariäärise defektid. • SV imendumine: toimub peamiselt soolelimaskesta hattude tipus. Monooside rohkuse korral toimub väikese koguse resorbeerumine ka passiivtranspordina. Imendumine on suht kiire ja peensoole algusosas praktiliselt lõppenud. Imendunud monoosid satuvad värativeeni kaudu maksa.
Süsivesikute metabolismi põhifunktsioonid. • SV katabolism peab andma 50-60% organismi toiduenergia vajadusest • Tagab veresuhkru taseme hoidmise normi piires • Tekitab monosahhariidseid eelühendeid • Häired selles avalduvad haigustena (suhkrutõbi ja rasvumine) Miks glükoos on inimkeha keskne süsivesik? • Lahustub väga hästi vees ja tema struktuur on optimaalse stabiilsusega • Vaba glükoos on organismis keemiliselt suht inertne (tema muundumine toimub aint ensümaatiliselt ja on seega täpselt kontrollitud) • Ta on metaboolne põhikütus enamike organismide jaoks • Läbib piisava kiirusega hemato-entsefaalset barjääri • Praktiliselt ainuke kütus ajukoe, arütrotsüütide, neerupealiste, silma võrkkesta ja testiste jaoks.
Veresuhkru mõiste. Veresuhkru hormonaalne regulatsioon • Veresuhkur on vere glükoosisisaldus. Glükoosi on veres söömata oleku ajal 3,4-5,2 mmol/l. Hüpoglükeemiaga on tegemist, kui veres on glükoosisisaldus normaalsest väiksem ja hüperglükeemiaga, kui normaalsest rohkem Vere glükoosisisaldust tõstavad kilpnäärme türoksiin (kiirendab glükoosi imendumist) ja adenohüpofüüsi kasvuhormoon (stimuleerib maksa glükoosisünteesi).Glükogeeni lõhustamist suurendavad glükagoon ja adrenaliin. Glükoneogeneesi reguleerivad neerupealise glükokortikoidid. Veresuhkru taset langetab insuliin.
Kuidas glükoos võetakse rakkudesse? Kuidas ta lülitub metaboolsetesse radadesse? Transpordiks läbi membraani on vajalikud valktransporterid (GLUT). GLUT2 transpordib glükoosi läbi enterotsüütide verre. GLUT1 ja GLUT3 viivad glükoosi rakku. GLUT2 võtab glükoosi maksarakkudesse. GLUT4 on insuliin-tundlik. Glükoos viiakse rakku Na-sõltuva ko-transpordiga Metaboolsetesse radadesse lülitub aktiveeritud vorm. Aktiveeritakse ATP-ga fosforüülides, glükoos-6-fosfaadiks
Anaeroobne glükolüüs Sellel on kaks faasi: esimeses toimub energia investeerimine, teises Glc lõhustumise energiamuudu osaline konverteerimine ATP-ks. Energeetiline kasum on 2 ATP.Anaeroobse glükolüüsi efektiivsus on 33%. Anaeroobse glükolüüsi regulatsioon Kudede glükolüütiline aktiivsus sõltub füsioloogilisest seisundist. Glükolüüsi kiir-kontroll toimub regulatoorsete võtmeensüümide aktiivsuse allosteerilise regulatsiooni kaudu. Üldmõjurid on metaboliitide (ATP, ADP, AMP, NAD, NADH) tasemed. Muutus nende norm kontsentratsioonides muudab regulatoorsete ensüümide aktiivsust. Reguleerijad: ATP muutub teatud tasemest alates signaaliks, et teda pole vaja enam toota, seega inhibeerib. • AMP taseme tõus tähendab aga ATP (energia) vähenemist, mistõttu annab aktiveeriva signaali. • Tsitraadi taseme kõrgenemine on signaaliks, et metaboliite ja energiat on piisavalt. • Vesinikioonid tekivad tänu laktaadi tekkele. • Fru-2,6,bisP on võimsam aktivaator. Teda tekitab Fru-6-P-st (kui selle tase tõuseb) FFK-2 kinaas-domeen. Kui Fru-2,6,bisP on palju lõhustab fosfataas-domeen ta tagasi Fru-6-P. Püruvaadi kinaasi regulatsioon: aktiveerib Fru-1,6,bisP ja AMP. Inhibeerib ATP
Anaeroobse glükolüüsi biokeemilis-meditsiiniline tähtsus • Eluliselt vaja tootmaks ATP anaerobioosis • Võimaldab elulemuse hüpoksia puhul. • Sobib väga järsult suurenenud energiavajaduse lühiajaliseks katmiseks (intensiivset tööd alustava skeletilihase rakkudesse ei jõua hapnikku piisavalt) • Tekkiv laktaat on glükoosi sünteesi reserv • Erütrotsüütides pole mitokondreid, niiet energiavajaduse katab anaeroobne glükolüüs • Aitab püruvaadi kuhjumist vältida. Selle kuhjumine võib põhjustada neurotoksilisi efekte
Laktatsidoos • Laktaadi tavahulgad venoosses veres on 0,5-2,2 mmol/l ja arteriaalses 0,3-0,8 mmol/l. • Kui laktaadi konts. vereplasmas on püsivalt üle 5mmol/l (hüperlakteemia) ja arteriaalse vere pH samaaegselt alla 7,25 on tegemist laktatsidoosiga. • Laktatsidoosi tavapõhjus on kudede hüpoksia, mitteadekvaatne vereringlus , samuti kui laktaadi kasutamine glükoneogeneesis on häiritud (maksas, neerudes) • Äge kopsuhaigus, uppumine, kõrgmäestik – hapnikuga varustamise häired • Leukeemia, kartsinoom – neoplastiliste rakkude AAG • Füüsiline tugev koormus – lihasrakkude AAG • CO mürgistus – hapniku transpordi häired • Tsüaniidi/alkoholi mürgistus – hingamisahela/glükogeneesi pärssimine
Aeroobne glükolüüs. Miks TKT on keskne metaboolne tsükkel? Aeroobne glükolüüs võimaldab SV täielikku oksüdatiivset lõhustamist. See lubab toota oluliselt suurema arvu ATP molekule kui anaerobioosis. Aerobioosis lõhustub glükoos püruvaadiks, mismuundatakse AcCoA-ks. See kondenseerub oksaloatsetaadiga ja siseneb TKT-sse. ´ TKT ühe ringi energeetiline kogusaagis on 12 ATP. TKT-sse kanaliseerub kogu metabolism. Ta seostab süsivesikute, lipiidide, aminohapete metabolismi. TKT põhiroll on AcCoA täielik lõhustumine süsihappegaasiks ja veeks ning lõhustumise energiamuudu arvelt rohke ATP tootmine. See toodab lõviosa ATP-st.
TKT regulatoorsed ensüümid. TKT talitluse häired Esüümid: • Tsitraadi süntaas ,akonitaas , isotsitraat. Allosteeriliselt inhibeerib seda ATP, NADH, aktiveerib ADP kõrgenev tase. • AKGDH – 3 ensüümi ja 5 vitamiinse koensüümi kompleks (tiamiin, riboflaviin, pantoteenhape, nikotiinhape, lipoehape). ATP, NADH, GTP ja suktsinüül-CoA tõus pärsib AKGDH tööd • Suktsinüül-CoA süntetaas, suktsinaadi degüdrogenaas (sisaldab raud-väävel tsentrit), fumaraas, malaadi dehüdrogenaas. Talitluse häired:On eriti ohtlikud intensiivse aeroobse metabolismiga kudedes. Hapniku vähenemisel langeb glükoosi aeroobne metaboliseerumine. Anaeroobne katabolism intensiivistub ja laktaadi kuhjumine tekitab laktatsidoosi, mis viib tasapisi ATP defitsiidile
TKT põhifunktsioon TKT on rakkude energiavajaduse põhitagaja. Tema vaheühenditest algab mitmeid biosünteese. TKT komponente kasutab rasvhapete ja steroolide, heemi, glükoosi süntees, mõndade aminohapete süntees. TKT-sse kanaliseerub kogu metabolism. Ta seostab süsivesikute, lipiidide, aminohapete metabolismi. TKT põhiroll on AcCoA täielik lõhustumine süsihappegaasiks ja veeks ning lõhustumise energiamuudu arvelt rohke ATP tootmine. See toodab lõviosa ATP-st.
Glükoneogeneesi vajalikkus • Glükoneogenees on glükoosi süntees mittesahhariidsetest ühenditest (laktaat, püruvaat, glütserool, AH). See toimub maksas, 10% ka neerukoores. See on vajalik hüpoglükeemia ära hoidmiseks. Kestval nälgimisel on glükoneogenees ainus variant hüpoglügeemia vältimiseks (Veresuhkru tasemel hoidmiseks) Üks glükoneogeneesi põhisubstraate on laktaat.
Glükoneogeneesi ja glükolüüsi vastastikune regulatsioon • Pärast toitumist tõusnud vere insuliin intensiivistab glükolüüsi ja pärsib glükoneogeneesi. Glükoosi kulutamiseks stimuleerib insuliin glükoosi kinaasi, Pyr kinaasi , FFK-1 ekspressiooni. • Toitumispausil ja nälgimisel tõuseb vere glükagoontase, mis annab maksale glükoneogeneesi intensiivistamise signaali. Soodustatud on PEP karbokinaasi, Glc-6-fosfataasi ja aminotransferaaside ekspressioon. Samuti tõstab glükagoon cAMP taset, mis tingib vastupidist reaktsiooni. • Glükagoon stimuleerib glükoneogeneesi ka lipolüüsi kaudu.
Glükogenolüüsi biokeemiline mehhanism • Algab ahelate lühendamisega, lühendatakse korraga paljusid ahelaid. Glükoosijääke eemaldab fosforülüütiliselt glükogeeni fosforülaas, glükogeeni ahela mitteredutseerivast osast Glc-1-P vormis. Eemaldamine kestab, kuni hargnemiskohani jääb 4 glükoosijääki. • Järgneb hargnemispunkti kõrvaldamine, seda teeb hargnemistkõrvaldav ensüüm glükosüül(4:4)transferaas. Transferaasse aktiivsusega võtab ta hargnemispunkti juurest fragmendi ja liidab selle lineaarahelale. Glükosidaasse aktiivsusega lõhub ta 1,6 sideme ehk hargnemispunkti, kust eraldub vaba glükoos. Nüüd on ahel glükogeeni fosforülaasi toimel edasi lõhustatav kuni järgmise hargnemispunktini. • Ennem tekkinud Glc-1-P muudab fosfoglükomutaas Glc-6-P-ks (millest nt maksas tehakse glükoos)
Glükogeneesi ja glükogenolüüsi allosteeriline regulatsioon • Normaalse toitumise järgne seisund ja puhkav lihas: substraatide saadavus ja metaboolse energia tase on kõrge. Glükoosi, Glc-6-P ja ATP taseme tõus inhibeerib allosteeriliselt maksa glükogeeni fosforülaasi (glükogenolüüsi) (lihastes inhib ka kreatiinfosfaadi tõus). Samal ajal aktiveerib Glc-6-P maksas allosteeriliselt glükogeneesi, kuna liigne glükoos tuleb salvestada. Reguleerija on ka AMP, mille madal tase näitab energiarikkust ja kõrge tase energiavajadust Intensiivselt töötav lihas ja paastumine/nälgimine: glükoosi, ATP, Glc-6-P ja kreatiinfosfaadi tasemed langevad oluliselt. Allosteeriline inhibitsioon lakkab, glükogeeni fosforülaas läheb aktiivvormi ja glükogeeni lõhustumine soodustub. Energiavajadusel AMP tase tõuseb, ta seostub inaktiivse glükogeeni fosforülaasiga ja aktiveerib selle (glükogenolüüs).
Glükogeneesi ja glükogenolüüsi hormonaalne regulatsioon • Glükogeeni fosforülaas ja glükogeeni süntaas esinevad aktiivvormis ja inaktiivses vormis. Normaalse toitumise järgne seisund ja puhkav lihas: toitumisjärgne veresuhkru tase põhjustab insuliini sekreteerimist. Tõusev insuliini tase lülitab maksa ja lihased glükogeeni sünteesile. Insuliin aktiveerib proteiin fosfataasi 1 (PF-1), mis omakorda aktiveerib glükogeeni süntaasi. Intensiivselt töötav lihas ja paastumine/nälgimine: substraatide saadavus on limiteeritud ja metaboolse energia tase madal. Seega on vaja hakata lõhustama glükogeeni maksas veresuhkru hoidmiseks ja lihastes kontraktsioonienergia tarbeks. Veresuhkru madalale tasemele reageerib pankreas glükagooni sekreteerimisega, mille seostumine retseptoritega lülitab maksa glükogenolüüsile (samal ajal stimuleerib ka glükoneogeneesi) ning samas inaktiveerib glükogeeni süntaasi. Adrenaliin aktiveerib glükogenolüüsi ja pärsib glükogeneesi. Toimib maksas ja lihastes ja tagab kiire ning rohke glükoosi mobilisatsiooni ja lõhustamise. Adrenaliin vabaneb vastuseks KNS stress-signaalile.
Pentoosfosfaadi tsükkel. • PFT on aeroobse glükolüüsi rada, mis toodab pentoosfosfaate ja NADPH. Ta toodab pea poole vajatavast redutseerivast energiast (NADPH). Kui on vaja riboos-5-P ja NADPH, siis suureneb Glc-6-P vool läbi PFT: • Toimivad nii intensiivsed taandavad sünteesid kui ka nukleotiidide sünteesid. Need rakud vajavad rohkesti NADPH ja riboos-5-P. Seega on intensiivistunud PFT esimesed reaktsioonid, teised limiteeritud. • Intensiivne rasvhapete süntees rasvkoes ja lakteeriva piimanäärme rakkudes, intensiivne steroidide süntees maksas ja neerupealistes. Seal vajatakse NADPH tohkem ja riboos-5-P vähem. Soodustatud on PFT esimene pool. • Intensiivne nukleiinhapete süntees. Vajatakse rohkem Rib-5P. Domineeriv on PFT teine pool ja esimene limiteeritud • Intensiivsed taandavad sünteesid. Sellised rakud vajavad NADPH ja ATP rohkem kui Rib-5P.
PFT ja NADPH PFT on NADPH põhitootja ja see kasutub: • Rasvhapete, steroidide sünteesiks • Ksenobiootikumide biotransformatsioonis maksarakkudes • Antioksüdantses võrgustikus – nimelt selle kaitsesüsteemi osa on GSH, mille koostöö GSH peroksüdaasiga elimineerib liigsed peroksiidid. Tekkiba oksüdeeritud glutatiooni (GSSG) viib kiiresti tagasi redutseeritud vormi GSH reduktaas NADPH abil. • Üldiselt on GSH paljudes kohtades vaja, ja tekkiva GSSG viib GSH reduktaas õigesse vormi tagasi NADPH abil. • Lämmastikoksiidi sünteesil • Fagotsütoosis – NADPH ja NADPH oksüdaasi koostöös tekib superoksiidi radikaal
Fruktoos ja kliinilised probleemid • Fruktoos leiab kasutamist aminosahhariidide ja siaalhapete sünteesis ja tal on ka mitmeid spetsrolle inimkehas. Tema sisenemine rakkudesse pole insuliin-sõltuv, niiet ta ei indutseeri insuliini produktsiooni eriti. Enamik toidufruktoosi töödeldakse maksas Fru kinaasiga Fru-1-P-ks, mis kasutub glükoneogeneesis, lipiidide sünteesis ja ka glükolüüsis. Lihastes tekib fruktoosist Fru-6-P heksoosi kinaasi abil. Kliinilised probleemid kestval liigtarbimisel: fruktoosi metabolism on kiirem kui glükoosi oma ja muutub takistamatult atsetüül-CoA-ks, mis intensiivistab lipiidide, rasvhapete sünteesi ja VLDL sekretsiooni maksas. Tekib hüperlipideemia, hüperkolesteroleemia.
Bioloogiline oksüdatsioon. Hingamisahela lihtsustatud skeem • Bioloogiline oksüdatsioon on elektronide transportahelate (ETA) põhine. Inimkeha rakkudes on kaks ETA põhitüüpi: hingamisahel ja kehavõõraste ühendite biotransformatsiooniga seotud ETA. Bioloogiline oksüdatsioon realiseerub redokspaaride (elektronide doonor/akseptor) töö kaudu. Süsteemide ensüümid on oksüdoreduktaasid, mis liigendatakse: • Oksüdaasid • Oksügenaasid • Hüdroksüperoksüdaasid • Dehüdrogenaasid Hingamisahelas elektronide plekanne toimub redokspaaride vahel, mis redokspotentsiaalide alusel reastuvad hingamisahelaks. Elektronid liiguvad negatiivsema RP redokspaarilt positiivsema RP redokspaarile, kujuneb kindel järjestus. Hingamisahel algab metaboliidilt vesinikuaatomite äravõtmisega ja lõpeb vee tekkega. Keskne komponend on koensüüm Q.