Download

1 / 62
660 likes | 765 Views
Hematopoiesis. Dr. Mervat Khorshied Ass. Prof. of Clinical and Chemical Pathology 2012/2013. Agenda. What is ment by haematopoiesis. Sites of haematopoiesis. BM microenvironment. Haematopoietic stem/Progenitor cells. Regulation of haematopoiesis. What is ment by Haematopoiesis ?

E N D
Hematopoiesis Dr. Mervat Khorshied Ass. Prof. of Clinical and Chemical Pathology 2012/2013
Agenda • What is ment by haematopoiesis. • Sites of haematopoiesis. • BM microenvironment. • Haematopoietic stem/Progenitor cells. • Regulation of haematopoiesis.
What is ment by Haematopoiesis? Is the process involved in the production of all blood cells from the pluripotent stem cell; The hematopoietic stem cells (HSC).
Hematopoietic organs • In the first few weeks of gestation, the yolk sac is the main site of haematopoiesis, however, definitive haematopoiesis derives from a population of stem cells first observed on the dorsal aorta termed the aorta-gonads-mesonephros (AGM) region. • These stem cells are termed Haemangioblasts as they are capable of differentiation into haematopoietic cells and endothelial cells.
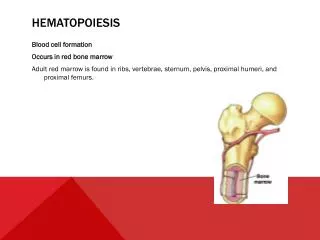
Fetal liver and spleen: from 6 weeks until 6 – 7 months of fetal life, and continue untill about 2 weeks after delivery. • Bone marrow: BM becomes the main site of hematopoiesis from the 6 – 7 ms of fetal life, which is distributed through the human skeleton but gradually recedes with time so that in normal adult life, hematopoiesis is found mainly in flat bones (sternum, pelvis, vertebrae, ribs and proximal ends of long femur.
Fetus: • Yolk sac: 0 – 2 ms • Liver and spleen: 2 - 7 ms • Bone marrow: 5 - 9 ms. • Infant: BM of all bones. • Gradual replacement of active (red) marrow by inactive (fatty) tissue. • Adults: Flat bones and proximal ends of femur
Bone marrow architecture/microenvironment:Bone marrow (BM) is a hematopoietic organ that resides within the protected confines of the bones and is the major location for hematopoiesis. In adults, active BM is distributed throughout the vertebrae, ribs, pelvis, skull and proximal ends of the long bones.
BM Structure:BM consists of a rigid bone cortex enclosing a cavity containing the arterial vascular system, a complex sinusoidal system, hematopoietic cells and the stroma. Arterial vessels enter the BM and divide into arterioles and capillaries that span throughout the BM to supply sinusoids, which in turn follows into the emissary vein and finally joins the systemic venous circulation.
Bone marrow stroma: • It is composed of stromal cells (fibroblasts, endothelial cells, adipocytes, macrophages) and a microvasculature network. Stromal cells secrete: • Extra-cellurar molecules as collagen, glycoproteins (fibronectin and thrombospondin) and glycosaminoglycans (hyaluronic acid and chondroitin derivatives) to form the extracellular matrix. • Several growth factors necessary for stem cell survival and function.
Stromal cells secrete extra-cellurar molecules as collagen, glycoproteins (fibronectin and thrombospondin) and glycosaminoglycans (hyaluronic acid and chondroitin derivatives) to form the extracellular matrix. • Additionally, stromal cells secrete several growth factors necessary for stem cell survival and function.
The BM sinusoids are unique, in that they are completely devoid of supporting connective tissue and, instead, the sinusoidal wall is made up of an overlapping single layer of endothelial cells. • Because of the lack of connective tissues, the major supporting structure for the sinusoids is the surrounding BM cells.
The cellular component of the BM can be divided into hematopoietic cells and mesenchymal-derived cells. • The mesenchymal-derived cells, along with the macrophages and the extracellular matrix (ECM), form the BM stroma. • The BM hematopoietic cells consist of hematopoietic stem cells (HSCs), hematopoietic progenitor cells (HPCs) and mature plasma cells. and HPCs
Interaction of stromal cells, growth factors and haemopoietic cells
- The HSCs and HPCs are held within the BM stroma not only through the adhesive interactions between VCAM-1, expressed by stromal cells, and integrin α4β1 expressed by the HSCs and HPCs, and by the chemotactic interaction between the chemokine CXCL12 and its receptor CXCR4 expressed on the HSCs and HPCs.- BM is also innervated by myelinated and unmyelinated nerve fibers.
Hematopoietic stem cell niche HSCs reside in the BM with a few circulating in the peripheral blood. HSCs are localized in a particular niche within the BM to maintain their self-renewal capacity. HSCs are localized at the bone–BM interface (Osteoblastic niche) and around the blood vessels (vascular niche). In contrast, the more committed progenitors accumulate in the center of the BM .
Osteoblastic niche The majority of HSCs in the endosteum are found to be in direct contact with the osteoblasts, and there is genetic evidence that increased bone formation leads to an increased number of HSCs in the BM. Several studies demonstrated the role of osteoblasts in the maintenance and survival of HSCs.
HSCs are held in the endosteum by the expression of adhesive molecules like osteopontin, N-cadherin, c-KIT ligand stem cell factor and hyaluronic acid. • In addition, the interaction is strengthened by the cytokines and chemokines that are produced by the osteoblasts, like chemokine CXCL12, angiopoietin-1, the ligand for the tyrosine kinase receptor Tie-2 expressed by HSCs, and jagged-1, a ligand for Notch1 receptor also expressed on HSCs
These interactions regulate HSC self-renewal by controlling the survival, quiescence and proliferation of HSCs.
Vascular niche: - Vascular niche was identified in the BM by using the signaling lymphocyte activation molecule (SLAM) family of receptors (CD150+CD48−CD41−). - SLAM family are cell-surface proteins that act as adhesion and signaling receptors and are differentially expressed among hematopoietic progenitors in a way that correlates with progenitor primitiveness.
One of the major physiological differences between the two niches is the high oxygen levels in the vascular niche compared to the hypoxic region of the osteoblastic niche. • The quiescent HSC located in the hypoxic region of the osteoblastic niche, on stimulation, could move to the vascular niche where the increased oxygen levels would induce the HSCs to undergo differentiation and provide the peripheral blood supply with the necessary mature hematopoietic cells.
When the differentiated hematopoietic cells are not required, HSCs move back to the endosteal niche and revert back to their original quiescent state, there by maintaining a constant number of differentiated hematopoietic cells in the peripheral blood .
Who are the HSCs? • HSC is the mother cell for all types of blood cells. It is a pluripotent cell capable of self renewal and differentiation. • The property of self renewal keeps the marrow cellularity (the stem cell pool) constant in a steady state in healthy individuals .
Self-renewal • Normally in G0 phase of cell cycle • The capacity for self-reproduction is vastly in excess of that required to maintain cell production for normal lifetime • As cells increase in number they differentiate as well • Multipotentiality • Capacity to generate cells of all the lymphohaemopoietic lineages
HSCs represents about 1 :20 million nucleated cells of the bone marrow. It resembles a small to medium sized lymphocyte. HSCs are capable to traffic around the body and are found in low number in the peripheral blood.
Stem cell division: Asymmetric division results in a stem cell dividing into either another stem cell or a more committed cell, while symmetric division yields either 2 stem cells or 2 differentiated daughter cells. This model predicts that cytokine-dependent self-renewal rate of hematopoietic stem cells is the most essential in hematopoiesis.
Progenitor cells • Encompasses from immediate progeny of stem cells to cells committed to one differentiation lineage • Progenitor cells become progressively more restricted in their differentiation and proliferation capacity. • Late progenitor cells eventually become restricted to one lineage.
One stem cell is capable of producing about 106 mature blood cells after 20 cell divisions.
Differentiation is the other fundamental property of hematopoietic progenitor cells and requires critical processes of cell-fate decision-making. • Decision making occurs as a result of biochemical signaling and gene regulatory networks within the cell. Ultimately, transcription factors determine cellular differentiation and specialization.
Regulation of Haemopoiesis Controlled cell production Controlled cell death There should be a balance between cell production and cell death except at the times of requirement
Haematopoietic growth factors • HGFs are glycoproteins that regulate the proliferation, differentiation of haematopoiuetic stem/progenitor cells and the functions of mature blood cells. • They may act locally at their site of production by cell to cell contact, or circulate in plasma. • They bind to the extracellular matrix to form niches to which stem and progenitor cells adhere.
They share a number of common properties: 1- Glycoproteins. 2- Produced by many cell types. 3- Active on stem/progenitor cells and on functional end cells. 4- Synergistic interactions. 5- Multiple actions: proliferation, differentiation, maturation , functional activation and prevention of apoptosis.
Growth factor receptors and signal transduction • HGFs mediate their actions through certain receptors on the target cells. • Most of HGFs receptors belong to the haematopoietin receptor superfamily , which dimerize after binding their ligand. • Dimerization of the receptor results in activation of a series of intracellular transcription pathways.
The three major pathways are:Janus associated kinase (JAK/STAT) pathway.The mitogen activated protein (MAP) kinase pathway.The phosphatidylinositol 3 (PI3) kinase pathway.
Binding of the GF to its receptor dimerization of these receptors Activation of tyrosine kinase domain generation of a cascade of biochemical events changes in gene expression, cell proliferation, differentiation, maturation or prevention of apoptosis.
A second smaller group of growth factors (GFs) as Stem cell factor (SCF) and macrophage colony stimulating factor (M-CSF) bind to another type of receptors with an extracellular immunoglobulin-like domain linked to the cytoplasmic transmembrane domain by a transmembrane brige.
Regulation and Requirements for Erythropoiesis The number of RBCs remains constant and reflects a balance between RBC production and destruction. Erythropoiesis is hormonally controlled and depends on adequate supplies of iron, amino acids, and B vitamins. Erythropoietin (EPO) is synthesized mainly from kidneys. Liver and BM macrophages may be also a source of Epo. Epo is secreted in response to anoxia caused for example by anemia or high altitude
Myelopoiesis Involves the action of a variety of growth factors e.g. stem cell factor (SCF), Fetal liver tyrosine kinase 3 (FLT3), Granulocyte Monocyte–Colony stimulating factor (GM-CSF), IL-3, Granulocyte–Colony stimulating factor (G-CSF), Monocyte–Colony stimulating factor M-CSF and IL-5.
Lymphopoiesis • Lymphocytes are immunologically competent cells. • Primary lymphoid organs: Bone marrow, Thymus. • Secondary lymphoid organs: Lymph nodes, Spleen, Lymphoid tissues. • Lymphocytes: • B and T lymphocytes • NK killer cells