Download
1 / 41
430 likes | 845 Views
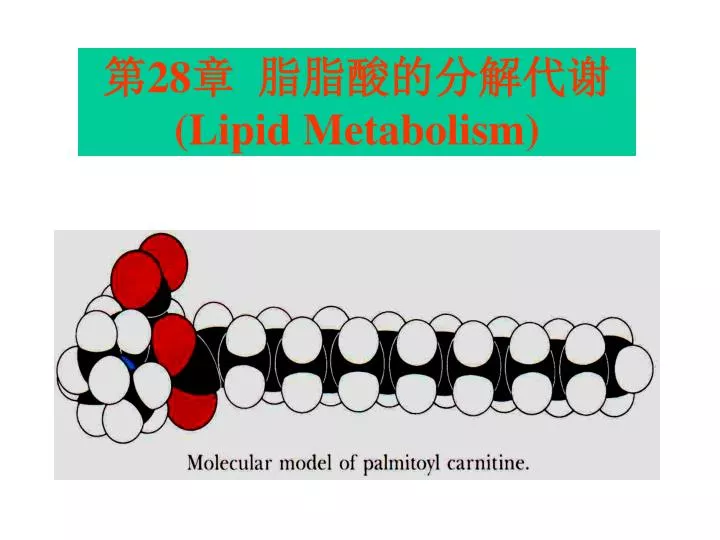
第 28 章 脂脂酸的分解代谢 (Lipid Metabolism). 脂质分类. 脂质基本都具有脂肪酸这一组成成份: 如:根据化学组成,脂质可分为: 1 .单纯脂质 ( 酯类) ( 1 )三酰甘油: 3 分子 脂肪酸 +1 分子甘油 ( 2 )蜡: 脂肪酸 + 长链醇或固醇 2 .复合脂质(酯类衍生物) ( 1 )磷脂: 甘油磷脂 和 鞘氨醇磷脂 ( 2 )糖脂: 甘油糖脂 和 鞘糖脂 鞘氨醇磷脂和鞘糖脂合称鞘脂类 3 .衍生脂质:取代烃,固醇类,萜,其它脂质. 甘油磷脂通式.

E N D
脂质分类 脂质基本都具有脂肪酸这一组成成份: 如:根据化学组成,脂质可分为: 1.单纯脂质(酯类) (1)三酰甘油:3分子脂肪酸+1分子甘油 (2)蜡:脂肪酸+长链醇或固醇 2.复合脂质(酯类衍生物) (1)磷脂:甘油磷脂和鞘氨醇磷脂 (2)糖脂:甘油糖脂和鞘糖脂 鞘氨醇磷脂和鞘糖脂合称鞘脂类 3.衍生脂质:取代烃,固醇类,萜,其它脂质
甘油磷脂通式 几种常见甘油磷脂极性头基 乙醇胺 胆碱 丝氨酸 肌醇
甘油糖脂 糖
鞘氨醇磷脂 鞘氨醇 神经酰胺 脂肪酸 磷酰胆碱
鞘糖脂 糖
脂类的分解代谢 一、脂肪的消化与吸收 二、脂肪酸的氧化(分解代谢) 三、酮体 掌握 四、磷脂的代谢(自学) 五、鞘脂类的代谢 六、甾醇的代谢 了解 七、脂肪酸代谢调节
脂肪酸功能及脂肪的贮能 脂肪酸具有多种生物学功能:(1) 组成成分;(2)蛋白膜定位导向作用;(3)能量贮存;(4)胞内信使。 P230。 关于能量方面:脂肪是非极性化合物,以无水状态存在,糖原是极性化合物,以水合状态存在,按同等重量计算,脂肪的代谢能量实际上高达糖原的6倍。
一、脂肪的消化和吸收 脂肪的消化开始于胃(胃脂肪酶)主要发生在小肠(胰脂肪酶): 1.胆汁盐乳化脂肪形成混合胶粒; 2.胰脂肪酶分解脂肪,最终分解为甘油和脂肪酸; 3.肠黏膜吸收分解产物甘油和脂肪酸,到体内再结合成脂肪;
脂肪的消化和吸收(续) 4.脂肪与胆固醇、蛋白质结合形成乳糜微粒; 5.乳糜微粒通过淋巴系统和血液循环进入 组织; 6.脂蛋白脂肪酶重新水解脂肪为脂肪酸和甘油; 7.脂肪酸进入细胞; 8.脂肪酸被氧化释放能量,或在肌细胞及脂肪组织中酯化储存。 9、甘油可转变为甘油-3-磷酸,再转变为二羟磷酸丙酮,进入氧化途径。
甘油进入酵解途径 P268 甘油激酶 甘油-3-P脱氢酶 磷酸丙糖异构酶
二、脂肪酸的氧化 Franz Knoop(1904)提出脂肪酸 -氧化假说,并通过苯基标记喂养试验,发现脂肪酸的氧化是从羧基端的位碳原子开始,每次分解出一个二碳片段(乙酰CoA)。 氧化发生在原核生物的细胞浆及真核生物线粒体基质。
(一)脂肪酸的活化 脂肪酸进入线粒体基质前首先被活化成脂酰 CoA。 酶:内质网脂酰CoA 合成酶,也称脂肪酸硫激酶。 反应需ATP Mg2+ R-COO-+ATP+HS-CoAR-CO-SCoA +AMP+PPi PPi2PI
(二)脂肪酸转入线粒体 脂肪酸(FA)的-氧化发生在肝脏及其他组织的线粒体内,中、短链FA(<10C)可直接穿过线粒体内膜,长链FA须经特殊的转运机制才可进入线粒体内被氧化,即肉碱转运机制。
肉碱与脂酰肉碱 肉碱 脂酰肉碱
肉碱转运 脂酰肉碱移位酶I 脂酰肉碱移位酶II
(三)脂肪酸的-氧化(主要) 包括四个反复氧化过程: 1. 脂酰CoA(n)的、脱氢,生成反式2烯酰CoA;——酶:脂酰CoA 脱氢酶; 2. 2烯酰CoA的水化,形成L-3-羟脂酰CoA,由水化酶催化,底物只能为2-烯酰CoA;——酶:烯酰CoA水合酶 3. L-3-羟脂酰CoA脱氢,生成-酮脂酰CoA,由脱氢酶催化,酶以NAD+为辅酶,只对L型底物有作用;——L-3-羟脂酰CoA脱氢酶 4.硫解(裂解、断链),生成脂酰CoA(n-2)由硫解酶(thiolase,acetyltransferase)(也叫酮脂酰硫解酶) 催化。 *四个反复的步骤:脱氢水化再脱氢硫解
脂肪酸-氧化 脂酰CoA 脱氢酶 烯酰CoA水合酶 L-3-羟脂酰 CoA脱氢酶 硫解酶
脂肪酸-氧化小结 1.FA仅需活化一次,消耗1ATP的两个高能磷酸键,活化的酶在线粒体膜外; 2.脂酰CoA(长链)需经肉碱运输才能进入线粒体内,有肉碱转移酶I和II; 3. -氧化的能量代谢,氧化产生的乙酰 CoA进入TCA,最终生成H2O和CO2,每一次循环产生1 分子乙酰 CoA、1FADH2和1(NADH+H+)。 以软脂酸为例,7次循环产生8 乙酰 CoA、7FADH2和7(NADH+H+) 总计:8?+7(1.5+2.5)-2=?(ATP)。
烃链含偶数C饱和脂肪酸的β-氧化 脂酰CoA 脱氢酶 烯酰CoA水合酶 L-3-羟脂酰 CoA脱氢酶 硫解酶
三、不饱和脂肪酸的氧化 (一)不饱和脂肪酸的氧化 不饱和脂肪酸同样需要活化和转运才能进入线粒体氧化,在遇到不饱和双键前进行常规的-氧化,若是顺式双键,必需经顺反异构为反式异构物,需两个酶:烯酰CoA异构酶,2,4-烯酰CoA还原酶。
9:不饱和双键在奇数 单不饱和脂肪酸的氧化 顺式 烯酰辅酶A异构酶 反式 *烯酰辅酶A异构酶具有两种作用:顺反异构转换,不饱和键转换
3:不饱和双键在奇数 不饱和脂肪酸的氧化 顺式 烯酰CoA异构酶 反式
4:不饱和双键在偶数 不饱和脂肪酸的氧化 脂酰CoA脱氢酶 2,4-烯酰CoA还原酶 顺式 烯酰CoA异构酶 反式
多不饱和脂肪酸的氧化举例 (1)试计算该图P241图28-13中不饱和脂酰辅酶A完全氧化净生成的ATP数目。 (2)试计算图28-12中不饱和脂酰辅酶A完全氧化净生成的ATP数目。
(二)奇数碳脂肪酸氧化生成丙酰-CoA 有少量的奇数碳脂肪酸,氧化时经多次-氧化最终产生若干乙酰辅酶A和一个丙酰CoA,丙酰CoA可生成琥珀酰CoA。 P242:图28-14生成琥珀酰CoA,消耗一个ATP。 *一分子的丙酰CoA完全氧化生成多少ATP?
丙酰-CoA的氧化代谢去路 *一分子的丙酰CoA完全氧化生成多少ATP?
脂肪酸的-氧化 脂肪酸-羟化酶
脂肪酸的-氧化:末端甲基氧化转变为二羧酸脂肪酸的-氧化:末端甲基氧化转变为二羧酸 单加氧酶 *鼠肝微粒体中存在,是一种少见的脂肪酸氧化途径, 该氧化方式加速脂肪酸降解,两端变成羧基后可同时进行-氧化
四、酮体 (一)乙酰CoA代谢结局 (1)进入TCA途径; (2)作为类固醇前体,用于合成胆固醇; (3)作为脂肪酸合成的前体; (4)在肝、肾、脑等组织中,尤其在饥饿、禁食、糖尿病等情形下,乙酰CoA可进一步缩合并生成乙酰乙酸、-羟丁酸和丙酮这三种物质,统称为酮体(ketone body)。
P244 (二)酮体合成 硫解酶 HMG CoA合成酶 -羟--甲基 戊二酰CoA HMG CoA裂解酶
酮体生成及发生酸中毒的原因 (1)饥饿和胰岛素水平过低会耗尽体内糖的贮存; (2)为获取能量,糖异生及脂肪酸氧化作用加速,导致草酰乙酸耗尽;
酮体生成及发生酸中毒的原因 (3)脂肪酸继续氧化,产生的乙酰CoA不能通过TCA被氧化,从而大量积累; (4)乙酰CoA→酮体。 酮体积累引起血液pH降低(酸中毒)、尿中酮体增高(酮病)。
乙酰乙酸 (三)肝外组织使用酮体作为燃料 琥珀酰辅酶A 琥珀酸 乙酰乙酰辅酶A 硫解酶 2 乙酰辅酶A
P245 -羟丁酸作为燃料
习题 • 肉毒碱 脂肪酸的ß-氧化 酮体 酮血症 • 脂酰辅酶A上每分解一分子乙酰辅酶A及其进入TCA循环彻底氧化,可产生ATP的分子数是: • A 15 B 16 C 17 • D 18 E 19 F 20 • 脂肪酸分解产生的乙酰CoA去路 • A、合成脂肪酸 B、氧化供能 C、合成酮体 • D、合成胆固醇 E、以上都是 • *脂肪酸的ß-氧化作用所必需的辅助因子有: • A. NAD+ B. NADP+ C. FAD • D. FMN E. HSCOA F. FH4 • *糖脂代谢中以FAD为辅基的脱氢酶有: • A.脂酰辅酶A脱氢酶 B.ß-羟脂酰辅酶A脱氢酶 • C.琥珀酸脱氢酶 D.苹果酸脱氢酶
习题 催化卵磷脂(磷脂酰胆碱)水解为磷脂酸和胆碱的酶是: A、磷脂酶A1B、磷脂酶A2 C、磷脂酶B D、磷脂酶C E、磷脂酶D 下列关于脂肪酸氧化的叙述除哪个外都是对的 A、脂肪酸过度氧化可导致酮体在血液中的含量升高。 B、脂肪酸的氧化需要肉毒碱作为载体。 C、脂肪酸的彻底氧化需要柠檬酸循环的参与。 D、脂肪酸进行β–氧化前的活化发生在线粒体内。 酮体在肝脏内产生,在肝外组织分解,酮体是脂肪酸彻底氧化的产物。 只有乙酰辅酶A是脂肪酸降解的最终产物。 脂肪酸经活化后进入线粒体内进行β-氧化,需经脱氢、脱水、加氢和硫解等四个过程。 在线粒体制剂中加入脂肪酸、CoA、O2、、ADP和Pi,可观察到脂肪酸的氧化,加入安密妥,十六碳脂肪酸彻底氧化为CO2和H2O,可生成多少ATP?为什么?