Download

1 / 29
290 likes | 483 Views
Traffic to and Function of Organelles A. Origins and characteristics of Organelles Overview B. Mitochondria & Chloroplasts Origins and characteristics Structure and function of Mitochondria Structure and function of Chloroplasts C. Peroxisomes Origins and characteristics

E N D
Traffic to and Function of Organelles • A. Origins and characteristics of Organelles Overview • B. Mitochondria & Chloroplasts Origins and characteristics Structure and function of Mitochondria Structure and function of Chloroplasts C. Peroxisomes Origins and characteristics Structure and function D. Apicoplasts Origins and characteristics Structure and function • E. Principles of Trafficking into Organelles • F. Trafficking into Mitochondria • G. Trafficking into Chloroplast • H. Peroxisomal import • I. Apicoplast trafficking • J. Comparison of trafficking in organelles J.R. Lingappa, Pabio 552, Lecture 5
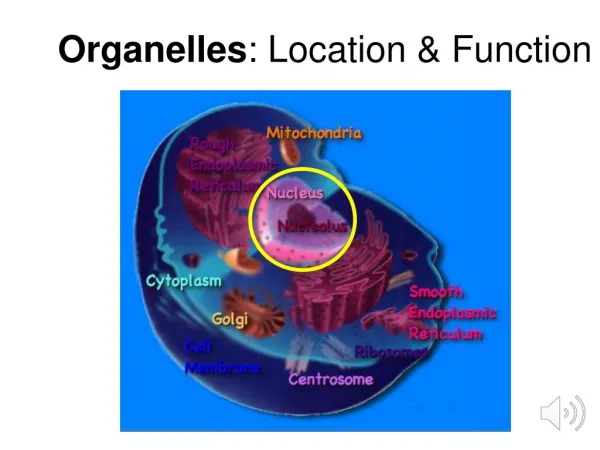
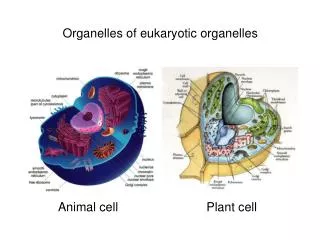
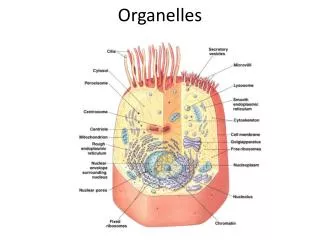
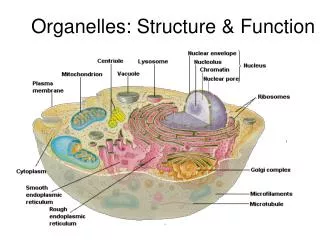
Trafficking to Organelles A. Origins and characteristics of Organelles: Overview 1. Organelles in all eukaryotes: Nucleus ER Golgi Lysosomes Endosomes Vesicles PM Mitochondria 2. Organelles in selected eukaryotes Plastids: Choloroplasts in plants Apicoplasts in toxoplasma and plasmodium (apicomplexans) Other secretory organelles: micronemes, rhoptries, dense granules in apicomplexans (see p. 25) Other unique organelles (see p. 26 ) J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles A. Origins and characteristics of Organelles: Overview 3. Evolution of Organelles: From Dyall et al. Science 304: 253, 2004 J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles B. Mitochondria (Mt) and Chloroplasts (Ch): 1. Origins and characteristics: a. Mt & Ch are organelles enclosed within a double membrane b. Contain their own genomes c. Arose symbiotically via engulfment of bacteria by ancestral eukaryotic cell (primary endosymbiosis). J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles B. Mitochondria (Mt) and Chloroplasts (Ch): 1. Origins and characteristics, cont: d. Most of their proteins are encoded in the nucleus (transfer of genetic responsibility to the host), translated free in the cytosol, & imported post- translationally in an unfolded state into Mt via specific targeting signals e. Some of their proteins are encoded by DNA in the organelle f. New Mt and Ch are formed by fission; cannot be produced de novo g. Contain ribosomes h. Transcription and translation occur in matrix I. N-formyl methionine as initiation codon just like in bacteria J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles B. Mitochondria (Mt) and Chloroplasts (Ch): Origins and characteristics, cont. How did genes get transferred from the endosymbiont to the nucleus? From Dyall et al. Science 304: 253, 2004 J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles • B. Mitochondria (Mt) and Chloroplasts (Ch): • 2. Mitochondrial Structure and Function: A. Structure: Outer membrane (OM; )Inner membrane (IM) has folds (cristae; Intermembrane space between IM and OM; Matrix is the interior. B. Function by compartment: 1. Matrix: Contains mitochondrial genome Encodes13 proteins (using a different genetic code), 2 rRNAs, 22 tRNAs Contains enzymes responsible for oxidative metabolism Oxidative metabolism: Conversion of glucose to pyruvate (glycolysis; anaerobic metabolism) occurs in cytosol; Pyruvate & fatty acids transported into Mt where they are converted to acetyl CoA, & oxidized to CO2 (citric acid cycle) to yield ATP, NADH, and FADH2 (aerobic metabolism). J.R. Lingappa, Pabio 552, Lecture 5
The Citric Acid Cycle Fatty Acid Metabolism Trafficking to Organelles • B. Mitochondria (Mt) and Chloroplasts (Ch): • 2. Mitochondrial Structure and Function: B. Function by compartment : 1. Matrix, cont. J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles • B. Mitochondria (Mt) and Chloroplasts (Ch): • 2. Mitochondrial Structure and Function: B. Function by compartment : 2. Membranes IM: NADH and FADH2 converted to ATP by oxidative phosphorylation; energy is stored in proton gradient in membrane impermeable to small ions and molecules. OM: Freely permeable to small molecules (<6kD) via porins that form channels Functions in Compartments J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles B. Mitochondria (Mt) & Chloroplasts (Ch): 2. Mt Structure and Function: B. Function by compartment, cont.: Mitochondrial proteins include proteins encoded in the nucleus and synthesized in the cytosol, as well as proteins encoded in the mitochondrion and synthesized in the mitochondrion. J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles B. Mitochondria (Mt) & Chloroplasts (Ch): 3. Chloroplast Structure and Function: A. Structure: OM, IM, and intermembrane space, and stroma (interior space), analogous to Mt Unlike Mt, Ch have an additional compartment (3rd membrane), the thylakoid. B. Functions: 1. Generation of ATP. 2. Photosynthetic conversion of CO2 to carbohydrates with production of O2. 3. Synthesis of amino acids, fatty acids, & lipid components of their own membranes. 4. Reduction of nitrate to ammonia. 5. Contains the Ch genome which encodes 120 genes & numerous rRNAs and tRNAs. J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles • Peroxisomes 1. Origins and characteristics: 2. Peroxisomes (P) are present in all eukaryotic cells, and: a. Differ from Mt because they are surrounded by only a single membrane, do not contain DNA or ribosomes, & acquire all their proteins by selective import from the cytosol b.Post-translational mechanism of protein import like that of the nucleus Does not involve unfolding of the cargo Involves a soluble receptor in the cytosol that recognizes a targeting signal Involves docking to proteins on the cytosolic surface of the peroxisome c.Resemble the ER: a single-membrane organelle replicating by fission d. Likely represent a vestige of an ancient organelle that performed all the oxygen metabolism of the primitive eukaryotic cell. Probably served to lower oxygen which was toxic to the primitive cell. Later, mitochondria developed and rendered peroxisomes somewhat obsolete because they carried out the same reactions but now coupled to ATP formation. J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles • Peroxisomes 2. Structure and Function: A. Structure: Organelle surrounded by a single membrane. B. Function (in animal cells): Contain peroxidases, which remove hydrogen ions from organic compounds, generating H2O2(hydrogen peroxide). RH2 + O2 = R + H2O2 Contain catalases, which use H2O2 to oxidize other substrates, including EtOH. H2O2 + R’H2 = R’ + 2H20 Oxidizes fatty acids, 2 carbons at a time, to acetyl CoA (occurs in mammalian Mt also). Formation of specific phospholipids found in myelin. J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles D. Apicoplasts 2. Origin and Characteristics: Apicoplasts (Ap) are homologues of chloroplasts, present in Apicomplexans (Plasmodium, Toxoplasma, Cryptosporidium): a.Complex plastids. b. Differ from Mt and Ch because: 1. Are surrounded by four membranes. 2. They originated from secondary endosymbiosis: primitive eukaryotic ancestor cell engulfed another eukaryote (green alga) that already possessed a chloroplast. 3. Contain proteins that traffic to the apicoplast via the secretory pathway. 4. Apicoplast proteins require an ER signal sequence. c.Resemble Mt and Ch because they: 1. Have their own genome (35 kB). 2. Require a transit peptide signal for protein import. d. Apicoplasts are required for infectivity. e. May be excellent drug targets because they contain prokaryotic metabolic pathways reflecting their origins. J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles D. Apicoplasts 2. Structure and Function: a. Structure: Organelle surrounded by 4 membranes. b. Function: Only discovered in the 1990's, so they have not yet been well studied Similar complex plastids found in algae as well Don’t perform photosynthesis (no genes for this), despite plastid origin. May play other metabolic roles, i.e. AA & FA biosynthesis, starch storage Apicoplast (A) in Plasmodium within an infected erythrocyte; From van Dooren et al., Parasitology Today 16, 421 (2000). J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles E. Principles of Trafficking into Organelles: 1. Traffic into Mt, Ch, and Pe constitute separate trafficking routes in the cell: A. ER-Golgi-Lysosomes/PM B. Cytoplasm-Nucleus C. Cytoplasm-PM D. Cytoplasm-Mt (or Ch) E. Cytoplasm-Pe 2. Distinguish between: Co-translational translocation -- ER Post-translational translocation of folded proteins -- nucleus Post-translational translocation of unfolded proteins -- mitochondria 3. Distinguish between: Transmembrane transport: channel closed when not translocating -- ER, mitochondria, etc. Gated transport: diffusion vs. selective transport across an open pore -- nucleus Vesicular transport -- Golgi, lysosomes, endosomes, PM. J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles E. Principles of Trafficking into Organelles: • Translocation of nuclear-encoded proteins into Mt & Ch is typically post-translational. A. Note that a very similar post-translational mechanism can be used in the ER of yeast and at bacterial plasma membranes. J.R. Lingappa, Pabio 552, Lecture 5
Translocators in Mitochondrial Membrane: Trafficking to Organelles E. Principles of Trafficking into Organelles: 4. Translocation of nuclear-encoded proteins into Mt & Ch is typically post-translational. • In contrast to the co-translational trans-location that in the eukaryotic ER, post-translational translocation into MT/Ch requires: 1. that newly-synthesized Mt protein be kept unfolded before translocation 2. the presence of a Mt signal sequence (also called presequence) that directs the chain to the OM 3. protein translocators in organelle mb that allow translocation across membrane 4. proteins targeted to organelles with multiple membranes often encode a second signal (transit peptide) to allow transport across inner membrane J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles F. Mitochondrial import: 1. Signals and Translocators: a. Mt import signal (pre-sequence) is an amphipathic helix J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles F. Mitochondrial import: 1. Signals and Translocators, cont.: b. Presequence binds to receptor on Mt surface. c. Insertion into the TOM complex = translocator across the outer Mt mb. Used by all proteins imported into Mt; mediated by the presequence. d. Insertion into TIM complexes (22 & 23) = translocators across inner Mt mb. mediated by a second sorting signal located distal to the presequence and, in the case of transmembrane proteins, a stop-transfer signal. e. Presequence removed in the matrix (or the intermembrane space) by signal peptidase. f. Thus, Mt proteins cross both membranes, which become closely apposed, at once rather than one at a time. J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles F. Mitochondrial import: 1. Signals and Translocators, cont.: : g. OxA complex mediates insertion of proteins synthesized in Mt into IM. h. Also proteins that are to be inserted into the IM are sometimes first translocated into the matrix, have their pre-sequence cleaved, & then the 2nd signal acts as an N-terminal signal directing them to be re-inserted into the IM via the OxA complex, with a stop-transfer to hold them in a transmembrane orientation. J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles F. Mitochondrial import, cont.: 2. Chaperones act on both sides of the mitochondrial membrane during translocation: a. Hsp70 maintains newly-synthesized Mt protein in cytosol in unfolded state. Release of protein from Hsp70 requires ATP hydrolysis. b. Translocation through the TIM complex requires electrochemical H+ gradient maintained by pumping H+ ions from matrix to inter Mt membrane space, driven by electron transport in inner mitochondrial membrane. Thus, electron transport in inner Mt membrane not only is the source of most of the cell’s ATP, but also transport of Mt proteins through TIM complex. J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles F. Mitochondrial import: 2. Chaperones act on both sides of the Mt membrane during translocation, cont.: c. Another Hsp 70 is associated with the TIM complex and acts as a motor that drives import. d. The translocated Mt protein is then transferred to an Hsp 60 chaperone in the matrix, which promotes Mt protein folding (and also hydrolyzes ATP). Two different Models for how Mt hsp70 drives protein import into the Mt J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles G. Chloroplast import is analogous to Mt import, except: 1. GTP and ATP are used for energy at OM and IM. 2. Electrochemical gradient is present at the thylakoid membrane. 3. Translocation complex in OM is Toc; translocation complex in IM is Tic. 4. Transit peptide directs translocation across OM and IM, and is removed by cleavage in the stroma, exposing in some cases a second signal sequence which directs transport across the thylakoid membrane. 5. While the signal sequences for Mt and Ch resemble each other, since both occur in plant cells, they need to be different enough to direct specific targeting to the right compartment. J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles H. Peroxisomal import: 1. Uses a 3 aa signal (Ser-Lys-Leu). 2. Attachment of this signal on a cytosolic protein results in peroxisomal import. 3. Driven by ATP hydrolysis. 4. Peroxins are proteins that participate in peroxisomal import. 5. Unlike in the case of mitochondria or chloroplasts, peroxisomal proteins do not have to be unfolded to be transported. 6. A soluble import receptor binds the cargo in the cytosol and accompanies it into the peroxisomes. After cargo releases, the receptor cycles back to the cytosol. This implies that an export system exists, but this has yet to be found. J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles I. Apicoplast import: 1. Related to chloroplasts but surrounded by 4 mbs. 2. Evidence exists for a classical secretory system in apicoplasts. 3. However, additional organelles exist (micronemes, rhoptries, and dense granules, and PVM). Also BFA not effective. 4. Leader sequence contains signal peptide (SP) transit peptide (TP). SP targets proteins to secretory system; SP + TP targets to apicoplast. 5. Toxo and plasmodium leader sequences function interchangeably. Chloroplast TPs from plants can also substitute for apicoplast TP. 6. TIC and TOC homologues are in apicoplasts. 7. Unclear if apicoplast is proximal or distal to Golgi. Legend: (a) Translation of protein with signal peptide followed by (b) Co-translational insertion into first membrane via SP; Second membrane recognizes TP; (c) Another Toc complex may be present in final set of membranes, perhaps acting along with a Tic complex (d). From van Dooren et al., Parasit. Today 16, 421 (2000) J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles I. Apicoplast Import: Apicoplast targeting is only one of the trafficking complexities of Toxo: From Joiner and Roos, J. Cell Biol. 157: 557-563, 2002 J.R. Lingappa, Pabio 552, Lecture 5
Trafficking to Organelles I. Apicoplast Import: Apicoplast targeting is only one of the trafficking complexities of Plasmodium: From van Dooren et al., Parasitology Today 16, 421 (2000): J.R. Lingappa, Pabio 552, Lecture 5
Additional Reading (not required): Dyall SD, Brown MT, Johnson PJ. Ancient invasions: from endosymbionts to organelles.Science. 2004 Apr 9;304(5668):253-7. Review. Osteryoung KW, Nunnari J. The division of endosymbiotic organelles. Science. 2003 Dec 5;302(5651):1698-704. Review. Wiedemann N, Pfanner N, Chacinska A.Chaperoning through the mitochondrial intermembrane space.Mol Cell. 2006 Jan 20;21(2):145-8. Review. Wickner W, Schekman R.Protein translocation across biological membranes.Science. 2005 Dec 2;310(5753):1452-6. Review. Horrocks P, Muhia D. Pexel/VTS: a protein-export motif in erythrocytes infected with malaria parasites. Trends Parasitol. 2005 Sep;21(9):396-9. van Dooren GG, Waller RF, Joiner KA, Roos DS, McFadden GI.Traffic jams: protein transport in Plasmodium falciparum.Parasitol Today. 2000 Oct;16(10):421-7. Review. J.R. Lingappa, Pabio 552, Lecture 5