Download

1 / 30
300 likes | 341 Views
Learn about the biosynthesis of fatty acids, from substrate and regulatory enzymes to variations and pathways within organisms.

E N D
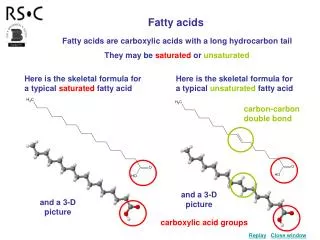
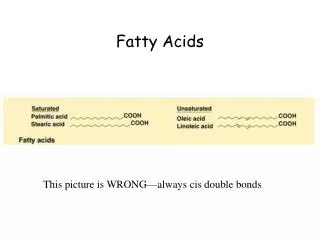
Fatty acids are a class of compounds containing a long hydrocarbon chain and a terminal carboxylate group.
Fatty Acid is synthesized from either carbohydrate or protein, when caloric intake exceeds daily requirements.
Fatty acid synthesis • substrate: acetyl-CoA, NADPH + H+ • product: palmitate (= endproduct of FA synthesis) • function: de novo synthesis of FA which are stored as TAG • subcelullar location: cytosol • organ location: mainly liver and adipose tissue and also other tissues (kidney, brain, lung, mammary gland) • regulatory enzyme: acetyl-CoA carboxylase
FATTY ACID SYNTHESIS • Variations in fatty acid synthesis between individuals may have a bearing on the nature and extent of obesity. Note :There is an inverse relationship between hepatic lipogenesis and the concentration of serum-free fatty acids.
Nutritional state regulateslipogenesis: • Lipogenesis converts surplus glucose and intermediates such as pyruvate, lactate, and acetyl-CoA to fat. • Rate is higher in well-fed animals whose diets contains a high proportions of carbohydrates. • It is depressed under conditions of restricted caloric intake, on a high-fat diet, or when there is a deficiency of insulin, as in diabetes mellitus. All these conditions are associated with increased concentrations of plasma free fatty acids.
Fat in the diet also causes depression of lipogenesis in the liver, and when there is more than 10% of fat in the diet, there is little conversion of dietary carbohydrates to fat.
Acetyl-CoA is formed from glucose via the oxidation of pyruvate and enters the mitochondria (glycolysis). However, it does not diffuse readily into the extramitochondrial cytosol the principal site of fatty acid synthesis. Citrate, formed after condensation of acetyl-CoA with oxaloacetate ( in the citric acid cycle within mitochondria), is translocated into the extramitochondrial compartment via the tricarboxylate transporter, ( T )
Citrate in the presence of CoA and ATP undergoes cleavage to acetyl-CoA and oxaloacetate catalyzed by ATP-citrate lyase, which increases in activity in the well-fed state. The acetyl-CoA is then available for malonyl-CoA formation and synthesis to palmitate
The resulting oxaloacetate can form malate via NADH-linked malate dehydrogenase, followed by the generation of NADPH via the malic enzyme. The NADPH becomes available for lipogenesis, and the pyruvate can be used to regenerate acetyl-CoA after transport into the mitochondrion. Note that the citrate (tricarboxylate) transporter in the mitochondrial membrane requires malate to exchange with citrate
This pathway is a means of transferring reducing equivalents from extramitochondrial NADH to NADP. Alternatively, malate itself can be transported into the mitochondrion, where it is able to re-form oxaloacetate.
Fat Synthesis Requires NADPH Malic enzyme provides ~ 50% of required NADPH. Remainder comes from the Pentose Phosphate Pathway.
Step 1: Formation of Malonylcoenzyme A is the committed step in fatty acid synthesis: It takes place in two steps: carboxylation of biotin (involving ATP) and transfer of the carboxyl to acetyl-CoAto form malonyl-CoA. Reaction is catalyzed by acetyl-CoA carboxylase. It is a multienzyme protein. The enzyme contains a variable number of identical subunits, each containing biotin, biotin carboxylase, biotin carboxyl carrier protein, and transcarboxylase, as well as a regulatory allosteric site.
The Formation of Malonyl CoA is the Committed Step in Fatty Acid Biosynthesis
Fatty Acid Synthesis • Overall goal is to attach a two-carbon acetate unit from malonyl-CoA to a growing chain and then reduce it • Reaction involves cycles of four enzyme-catalyzed steps • Condensation of the growing chain with activated acetate • Reduction of carbonyl to hydroxyl • Dehydration of alcohol to trans-alkene • Reduction of alkene to alkane • The growing chain is initially attached to the enzyme via a thioester linkage • During condensation, the growing chain is transferred to the acyl carrier protein • After the second reduction step, the elongated chain is transferred back to fatty acid synthase
Step 2: • FATTY ACID SYNTHASE catalyzes the remaining steps. • It is a multienzyme polypeptide complex that contains acyl carrier protein (ACP). ACP contains the vitamin pantothenic acid in the form of 4'-phosphopantetheine. ACP takes over the role of CoA. • Synthesis of all enzymes in the complex is coordinated, since it is encoded by a single gene • It is a dimer, and each monomer is identical, consisting of one chain containing all seven enzyme activities of fatty acid synthase and an ACP with a 4'-phosphopantetheine-SH group. Dimer is arranged in a "head to tail" configuration. Monomer is not active.
It is a dimer, and each monomer is identical, consisting of one chain containing all seven enzyme activities of fatty acid synthase and an ACP with a 4'-phosphopantetheine-SH group. Dimer is arranged in a "head to tail" configuration. Monomer is not active.
Enzymatic Activities in Fatty Acid Synthase • Condensationwith acetate • -ketoacyl-ACP synthase (KS) • Reductionof carbonyl to hydroxyl • -ketoacyl-ACP reductase (KR) • Dehydrationof alcohol to alkene • -hydroxyacyl-ACP dehydratase (DH) • Reduction of alkene to alkane • enoyl-ACP reductase (ER) • Chain transfer • Malonyl/acetyl-CoA ACP transferase
To start the second cycle, butyryl-ACP condenses with malonyl-ACP to form hexanoyl-ACP. • Overall goal is to attach a two-carbon acetate unit from malonyl-CoA to a growing chain and then reduce it • Reaction involves cycles of four enzyme-catalyzed steps • Condensation of the growing chain with activated acetate • Reduction of carbonyl to hydroxyl • Dehydration of alcohol to trans-alkene • Reduction of alkene to alkane • The growing chain is initially attached to the enzyme via a thioester linkage • During condensation, the growing chain is transferred to the acyl carrier protein • After the second reduction step, the elongated chain is transferred back to fatty acid synthase
To start the second cycle, butyryl-ACP condenses with malonyl-ACP to form hexanoyl-ACP.
To start the second cycle, butyryl-ACP condenses with malonyl-ACP to form hexanoyl-ACP. Note: Malonyl-ACP feeds in at each cycle. The same pattern continues until the product of the 7th cycle, palmitoyl-ACP undergoes hydrolysis to yield palmitate and free ACP.
REGULATION OF LIPOGENESIS SHORT TERM synthesis is controlled by allosteric and covalent modification of enzymes LONG TERMthere are changes in gene expression
Short-term • Acetyl-CoA carboxylaseis most important in regulating synthesis • Activated by citrate, which increases in well-fed state and is an indicator of a plentiful supply of acetyl-CoA • Inhibited by long-chain acyl-CoA. • Pyruvate dehydrogenaseregulates availability of free acetyl-CoA for lipogenesis. Acetyl-CoA causes an inhibition of pyruvate dehyrogenase.
Hormones (short term) • Insulin stimulates lipogenesis by several mechanisms: • a. increases transport of glucose into the cell (e.g., adipose tissues) and thereby increases the availability of both pyruvate for fatty acid synthesis and glycerol-3-phosphate for esterification of the newly formed fatty acids. • b. Converts inactive form of pyruvate dehydrogenase to the active form in adipose tissues • c. Activates acetyl-CoA carboxylase • d. Insulin inhibits lipolysis • e. Insulin antagonizes the actions of glucagon and epinephrine
Hormonal Regulation Phosphorylation inactivates (i.e., with low blood glucose, will breakdown fat, not synthesize it)