Download

1 / 54
540 likes | 701 Views
Non-conducting functions of voltage-gated ion channels. Leonard Kaczmarek Oktober 2006, nature review neuroscience. Einführung :. Grundlagen für das Verständnis leitender Funktionen von spannungsabhängigen Ionenkanälen. Klassifizierung von Ionenkanälen Gating Aktionspotential

E N D
Non-conducting functions ofvoltage-gated ion channels Leonard Kaczmarek Oktober 2006, nature review neuroscience
Einführung : Grundlagen für das Verständnis leitender Funktionen von spannungsabhängigen Ionenkanälen
Klassifizierung von Ionenkanälen • Gating • Aktionspotential • Alternatives Gating
Klassifizierung von Ionenkanälen Kationenkanäle: hohe Selektivität (Namen!) Ionenkanäle Anionenkanäle: geringe Selektivität spannungsabhängig (präsynaptisch) Gating ligandenabhängig (postsynaptisch)
Gating • 2 Zustände von Ionenkanälen: offen u. geschlossen • Energie für Konformationsänderung stammt bei den spannungsabh. Kanälen aus der Änderung der Membranspannung • Kaskade von Konformationsänderungen
Aktivierung <-> Deaktivierung • Übertragung der Energie auf Spannungssensor des Kanals • Bewegung des Sensors im elektrischen Feld • Konsequenz: weitere Konformationsänderungen im Kanal Resultat: Öffnung der Pore In Nav-Kanälen geschieht dies in weniger als einer Millisekunde, bei Kv-Kanälen dauert es deutlich länger: 10 bis ungefähr 70 ms.
Kanalinaktivierung (N-Typ) • bei Nav-Kanälen und bei einigen Kv-Kanälen (A-Typ-Kanäle) • führt zur Unterbrechung des Ionenstroms • Innerhalb weniger Millisekunden nach Aktivierung
Weitere Mechanismen C-Typ-Inaktivierung: • Beruht auf Konformationsänderungen im Bereich des Selektivitätsfilters • unabh. aber beschleunigt durch N-Typ-Inaktivierung • besonders langsame Rückreaktion
Ruhepotential • Nahe beim Gleichgewichtspotential für Kalium: -70 mV • Leitfähigkeit für Kalium über Kir-Kanäle oder 2-P-Domänen Kaliumkanäle sichergestellt
Aktionspotential I: Initiationsphase II: Depolarisation IIa: Aufstrich IIb: Overshoot III: Repolarisation IV: Nachhyperpolarisation Transiente Änderung des Membranpotentials, ausgelöst durch einen Reiz, der die Zelle über einen Schwellenwert hinaus depolarisiert.
Depolarisation • Stimulus depolarisiert Membranpotential bis zum Schwellenwert • Spannungsabhängige Natriumkanäle (Nav) öffnen sich, Na+ strömt in die Zelle • Kir-Kanäle (o. Ä.) werden inaktiviert
Repolarisation • Inaktivierung der Nav-Kanäle • Öffnen der Kv-Kanäle
Nachhyperpolarisation • Ca2+-abhängige Kaliumkanäle • Konzentration wird mit der Zeit gesenkt
alternatives Gating Ionenkanäle können durch versch. Signale geöffnet bzw. verschlossen werden: • intrazelluläre Messenger • Proteine • mechanische Spannung • Wärme/Kälte • kleinmolekulare Porenblocker
Intrazelluläre Messenger • ATP an 2-Segment-Kalium-Kanälen (A) • cAMP und cGMP an HCN- und CNG-Kanälen (B) Andere über: pH, oder Ca2+
HCN-Kanäle • nicht-selektive Kationenkanäle • verantwortlich für Ih-Strom • Langzeitbahnung der synapt. Antworten, Ausschüttung von Neurotransmittern über mehrere Stunden • Initiieren Langzeitbahnung
physikalische Faktoren • Umgebungsqualitäten (Kälte, Wäreme, mechanische Zugkraft und Osmolarität) können in Kanalgating umgesetzt werden. -> hierfür verantwortlich: TRP-Kanäle
Gliederung - Einführung: altes und neues Verständnis- Kaliumkanäle- Natriumkanäle- Calciumkanäle- TRP – Kanäle - Nichtleitende Funktionen von Ionenkanälen und Krankheiten - Fazit
Einführung: altes und neues Verständnis - primäre Funktion spannungsabhängiger Ionenkanäle: Erzeugung verschiedener Typen von Feuermustern für Informationsprozessierung und motorische Outputs - Modulierbarkeit (enzymatisch) ermöglicht Anpassung elek- trischer Eigenschaften des Neurons an die Inputsituation Diese Modulation wird üblicherweise als einseitiger Prozess betrachtet, bei welchem die Ionenkanäle als passive Elemente verändert werden
- aktuell (bis Okt. 2006) : Hinweise auf direkte Beeinflussung biochemischer Ereignisse in Zellen durch Ionenkanäle ohne (direkte) Notwendigkeit der leitenden Funktion als Ionenkanal - direkte Aktivierung von Enzymen intrazellulärer Signalwege- Funktion als Adhäsionsmoleküle oder Komponenten des Cytoskelettes- Regulation der Expression bestimmter Gene
methodische Grundlagen und Ergebnisse zitierter Studien : - Deletion (/Knockout) / Überexpression / Transfektion verändert Zelleigenschaften auf Arten, die nicht durch bloße Veränderung der Erregbarkeit erklärt werden können- Untersuchungen mit nichtleitenden Mutanten gleiche Effekte - vom Ionenfluss unabhängige Funktionen bestimmter Untereinheiten gefunden
Kaliumkanäle • erhöhte Expression / Aktivität in Tumorzellen Hinweis auf Zell-regulative Funktionen - Transfektion in Gliazellen / nicht- neuronale Zellen : Initiierung / Beschleunigung der Zellproliferation These: direkter Einfluss auf cytoplasmatische und nukleare Signalwege
Kv10.1 K+ - Kanal (EAG) - bisher als weitgehend typisch für synaptische Enden von Neuronen betrachtet - Drosophila melanogaster mit eag – knockout : Habituation und assoziatives Lernen verändert
Besondere Eigenschaften der cytoplasmatischen Domänen: - Drosophila: EAG bindet eine Kinase (CaMKII; α- UE), was zu deren konstitutiver Aktivierung führt - (C)- Terminus : nuclear localization signal- (N) – Terminus: PAS – Domäne (bei anderen Proteinen regulativer Einfluss auf Transkription und Proteinkinase Aktivität)
Beteiligung des Spannungssensors an regulativen Einflüssen: - Beeinflussung der Aktivität des Signalweges der Mitogen-aktivierten Proteinkinase (MAPK) - Transfektionsexperimente mit Fibro- / Myoblasten: Erhöhung von Zelldichte und DNA –Synthese unter gesteigerter Aktivität der p38 MAPK; unabhängig von der Leitfähigkeit des EAG, aber abhängig von Gating – Zustand des Kanals !
Einfluss des Gatingzustandes auf regulative Funktionen des EAG - Kanals - Verwendung von Mutanten, bei welchen die Spannungsab- hängigkeit der Aktivierung in den Bereich negativer Potentiale verlagert ist Kanal dauerhaft geöffnet - Folge : Blockade des proliferativen Effekts EAG Kanal muss sich zur Aktivierung der p38MAPK im geschlossenen Zustand befinden [Beleg für die Unabhängigkeit des Effekts vom Ionenfluss !]
Kv1.3 - Kanal - u.a. im Hippocampus und bulbus olfactorius exprimiert - besondere Eigenschaft: zeigt bei wiederholter Depolarisation kumulative Inaktivierung (K+ - Kanäle: Repolarisation ) zeitliche Ausdehnung von AP; erhöhte Erregbarkeit bei wiederholter Stimulation des Neurons
Effekte der Aktivität des KV1.3 – Kanals auf Zellproliferation -in Transfektionsexperimenten vom Ionenstrom durch den Kanal abhängig gewesen ! (Porenmutanten /K+-Ionophore) • In vivo – Hinweise : neben Beeinflussung des Membranpo- tentials und K+ - Strom weitere Aspekte relevant
Experimente mit Kv1.3 knockout - Mäusen - deutlich erhöhte Level von Signalmolekülen im bulbus olfactorius - niedrigere Konzentrationsschwelle für das Entdecken von Duftstoffen - erhöhte Fähigkeit zur Unterscheidung verschiedener Duftstoffe - diese Effekte traten nicht beim Eliminieren anderer K+ - Kanäle auf Ursache nicht auf bloße Veränderung der Membran- erregbarkeit beschränkt
möglicher Mechanismus des spannungsunabhängigen Effektes : - Kv1.3 in intakten Zellen eng mit β – Integrinen assoziiert - diese regulieren Zell –Zell- Interaktionen, Zellwachstum und -überleben - spezifische Kv1.3 – Blocker heben die Assoziation mit Integrinen auf Kv1.3 – Aktivität trägt vermutlich direkt zur Integrin – Regulation bei
Koordination neuronaler Erregbarkeit - Koordination der Expression von Genen für Kanäle bis dato ungeklärt aktuelle Studien : - Expressionslevel von Kv4 – Kanälen beeinflusst Expression eines anderen Kanals; dieser Effekt könnte vom Ionenfluss unabhängig sein - genauer Mechanismus noch unklar
Enzymatische Aktivität der β – Untereinheiten von Kv - Kanälen - neben Assoziation von Kinasen (EAG;α- UE) häufig Bindung zusätzlicher Untereinheiten gefunden - einige dieser zusätzlichen UE zeigen enzymatische Aktivität - möglicherweise vermitteln sie einige der Effekte von K+ - Kanälen auf zelluläre Signalwege
Bsp.: Familie des K+ - Channel interacting protein (KChIP1-4) - direkte Bindung an Kv4 α – UE, welche schnell inaktivierende A-Typ Kaliumauswärtsströme erzeugen Beeinflussung von Amplitude und Kinetik der Kv4 – Inaktivierung durch KChIP1-4 - ferner Funktion als Ca2+ -bindende Regulatoren von Presenilinen (membranintegrale Spaltenzyme) und Ca2+ - abhängige Transkriptionsfaktoren
Zusammenfassung: nichtleitende Funktionen von K+ - Kanälen - direkter Einfluss auf cytoplasmatische und nukleare Signalwege- Effekte auf Signalübertragung können z.B. vom Gatingzustand abhängen- Effekte beeinflussen perzeptuelle / kognitive Fähigkeiten durch Regulation verschiedener zellulärer Prozesse und können darüber hinaus Ausbruch von Krankheiten begünstigen
Na+ - Kanal β – Untereinheiten als Adhäsionsmoleküle - CAMs (Cell Adhesion Molecules) mit kurzem cytoplasma- tischem C – Terminus und langen extrazellulären Domänen • vermitteln Zell – Zell – Adhäsion durch homo- oder hetero- typische Interaktionen und Interaktionen mit Molekülen der extrazellulären Matrix
- an der cytoplasmatischen Seite Bindung an Proteine, welche ihrerseits mit dem Cytoskelett verbunden sind (Ankyrin G undB) β – Untereinheiten ermöglichen Na+ - Kanälen eine physikalische Verbindung des Cytoskeletts mit der Oberfläche von Nachbarzellen - vermutlich trägt diese Interaktion zur Ausbildung von Axonhügeln und Ranvier´schen Schnürringen bei (hohe Dichte von Na+ - Kanälen)
Einflüsse der Untereinheiten auf das Wachstum von Neuriten - β1 – Untereinheiten können das Wachstum von Neuriten bei dissoziierten Neuronen stimulieren - β2 – Untereinheiten dagegen können die Länge von Neuriten durch einen Effekt reduzieren, der durch α- oder β1- Unter- einheiten aufgehoben werden kann nicht- leitende Funktionen von Na+ - Kanälen könnten wichtige organisatorische Elemente der neuronalen Entwicklung sein
Regulation von β –Untereinheiten der Na+ - Kanäle via Spaltung - β –Untereinheiten werden kontinuierlich durch proteolytische Spaltung umgesetzt - möglicherweise reguliert Spaltung der extrazellulären und anschließend der (C) – terminalen Domäne die Beteiligung der β2 – Untereinheit an Prozessen der Zelladhäsion und -migration
Ca2+-Kanäle • Ca2+-Kanäle wohl am engsten verschränkt mit zellulären Ereignissen • Wie kann es sein, dass versch. Wege der Konzentrationserhöhung von Ca2+, verschiedene Effekte auf die Zelleigenschaften haben? • Beispiel: Aktivierung des Ca2+-Einstroms durch L-Typ Ca2+-Kanäle der Cav-Familie, aktiviert die MAPK-ERK-Signalkaskade. Dadurch wird CREB, ein Transkriptions-faktor aktiviert.
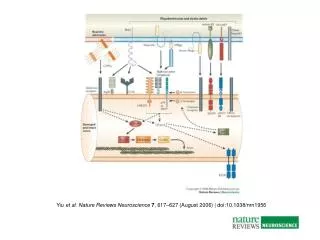
elektromech. Kopplung in Skelettmuskelzellen • Signal unabhängig vom Ionenfluss • phys. Verbindung des Ca2+-Kanals mit Ryanodin-Rezeptor -> Reaktion als Kanal, aber auch als Spannungssensor für Ryanodin-Rezeptor
außerdem: Verbindung zwischen Ca2+-Kanälen und dem Exocytose-Komplex • Interaktion mit SNARE-Proteinen -> enge Verbindung zwischen Vesikeln und den Kanälen
Hinweise, dass die physikalische Verbindung Informationen über den Zustand der Kanäle zur release site übertragen kann • Ausschüttung von Neurotransmittern unabhängig vom Ca2+-Einstrom • Depolarisation der Membran führt zur Konformationsänderung im Kanal und einer Änderung der Aktivität der SNARE-Proteine • Effekt kann verhindert werden durch intrazelluläre Blockade der Bindung zwischen Cav2.1-Kanälen und den SNARE-Proteinen
TRP-Kanäle • nicht-selektive Kationenkanäle • Einstellung des Ruhemembranpotentials, regulieren Konzentrationen von Ca2+ und Mg2+ in Neuronen und anderen Zellen • Überexpression in Krebszellen