Download
1 / 68
780 likes | 1.22k Views
Mutation and DNA Repair. Mutation Rates Vary Depending on Functional Constraints. Low Mutation Rates are Necessary for the Evolution of Complexity.

E N D
Low Mutation Rates are Necessary for the Evolution of Complexity 1. Because most mutations are deleterious, there are limits to the number of mutations that an organism can afford to accumulate in its somatic body, e.g., a) given mean eukaryotic rates, genomes can accommodate 60,000 genes without intolerable mutational loads (Alberts et al.) b) a mutation rate 10 times higher would limit genome size to ca. 6000 genes 2. Both the germ line and the somatic body must be protected from mutational load (rare mutations become common because of large genomes and cell proliferation), e.g., a) germ line: (1) DNA repair (2) meiotic recombination in all eukaryotes (3) sequestering of germ line in metazoans (4) diplontic selection among cell lineages in meristems of plants b) somatic tissues ...20% of deaths in western societies are due to cancer (uncontrolled cell proliferation) resulting largely to the accumulation of genetic damage in somatic tissues (1) DNA repair (2) immune systems 3. less efficient DNA repair and absence of meiosis may explain the limitation of prokaryotes to small genomes and unicellular forms before the origin of these processes in the protoeukaruote line. 4. spontaneous nucleotide changes are much higher than mutation rates would indicate, because of DNA repair mechanisms
Genotype to Phenotype - sequencing and searching for homologous sequences, then study their function Phenotype to Genotype - mutational screens and functional analysis Two strategies to study gene function
Point Mutations Same sense mutations Missense Mutations Nonsense Mutations Transitions Transversions Frame shift mutations Substitutions, Deletions and Additions Kinds of Mutations
Chemistry of single nucleotide substitutions: a) transitions: a pyrimidine replaces a pyrimidine (C T or T C) or a purine replaces a purine (A G or G A) b) transversions: a pyrimidine replaces a purine or vice versa c) transitions are less severe mutations that transversions: (1) chemically, purines are more similar to one another than they are to pyrmidines, and vice versa (2) genetically, amino acid substitution is less likely with transitions because of the degeneracy of the genetic code (a) 3rd position transitions often code same amino acid i) UUU and UUG both code for leucine ii) GAA and GAG both code for glutamic acid (b) 3rd position transversion less often codes for same amino acid i) UUU and UUG code for phenylanaline and leucine
Mutagenesis • Spontaneous Mutations • Replication Errors • Other Errors • Chemical Mutagenesis • Radiation-induced Mutations
Mutator Strains of E. coli • error prone replication • mutD codes for e subunit of DNA pol III:
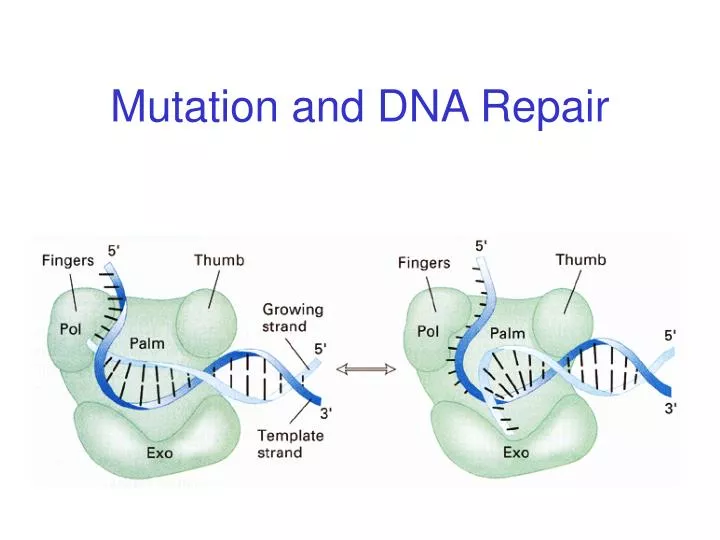
DNA polymerase III holoenzyme with subunits (weight in daltons) e Step 1: previous nucleotide pair is tested for complementarity. If passed, elongation occurs. Step 2: If failed, the elongating strand is transferred to the exonuclease site to excise the mismatched nucleotide.
Experimental Demonstration of Proofreading last nucleotide is non-complementary and labeled double labeled probe artificial template non-complementary nucleotide excised, but no complementary nucleotides
AT GC AT AT replication replication replication replication Replication A • Tketo template daughter AT replication AT A • Tketo replication replication A • Tketo daughter template AT replication AT replication A • Tketo template daughter A • Tenol replication if unrepaired G • Tenol daughter template
GC AT AT AT replication replication replication replication Replication Aamino • T template daughter AT replication AT Aamino • T replication replication Aamino • T daughter template AT replication AT replication Aimino • C template daughter if unrepaired Aimino • T replication Aamino • T daughter template
Cytosine Tautomers : Camino•G Cimino•A bindingmutation fromC to T common results in C•G pairing rare results in C•A pairing AT substitution
TA CG CG CG replication replication replication replication Replication Camino • G template daughter CG replication CG Camino • G replication replication Camino • G daughter template CG replication CG replication Cimino • A template daughter if unrepaired Cimino • G replication Camino • G daughter template
Guanine Tautomers : Gketo•C Genol•T binding mutation fromG to A common results in G•C pairing rare results in G•T pairing
TA GC GC GC replication replication replication replication Replication Gketo • C template daughter GC replication GC Gketo • C replication replication Gketo • C daughter template GC replication GC replication Genol • T template daughter if unrepaired Genol • C replication Gketo • C daughter template
Frameshift Mutations insertion
Mechanism of Frameshift Mutation: “Slipping a cog” …a base fails to pair with its partner during replication
Spontaneous hydrolysis can result in deamination and depurination
Deaminationreplacement of an amino group by a carbonyl oxygen These nucleotide analogs have different pairing affinities, but analogs can be recognized and repaired 5-methyl C deamination results in T, which can’t be recognized as a mutation Replication produces a GC and an AT C’s are selected for methylation in certain CG sequences, which has led to the conversion of most CG’s to TG’s during evolution
Deamination of C and Aillustrating different pairing behavior
Deamination and repair of C Deamination
Amplification of copy number by unequal crossing-over Unequal crossing-over becomes more likely with increased copy number
Dynamic Mutations Unequal crossing-over becomes more likely with increased copy number and The severity of the pathology increases with copy number therefore... Both the probability of the pathology and its severity increase over generations after the number of repeats approaches the threshold
A number of conditions are based on this mechanism operating in different genes
The repeats can be located in different orientations with regard to the coding sequence upstream downstream within within
The repeats can be located in different orientations with regard to the coding sequence ...even within a single gene
Nucleoside analogs can exhibit variant pairing behavior keto (above); enol pairs to G instead of A
Acridine dyes intercalate DNA sequences Effect: stabilizes the looping that leads to deletions and insertions that cause frame shift mutations
Mechanism of Frameshift Mutation: “Slipping a cog” …a base fails to pair with its partner during replication
Major Repair Mechanisms • Mismatch repair • Excision repair • Double strand breaks repaired mainly by end-joining • Inducible & error-prone mechanisms






















