Download
1 / 48
480 likes | 712 Views
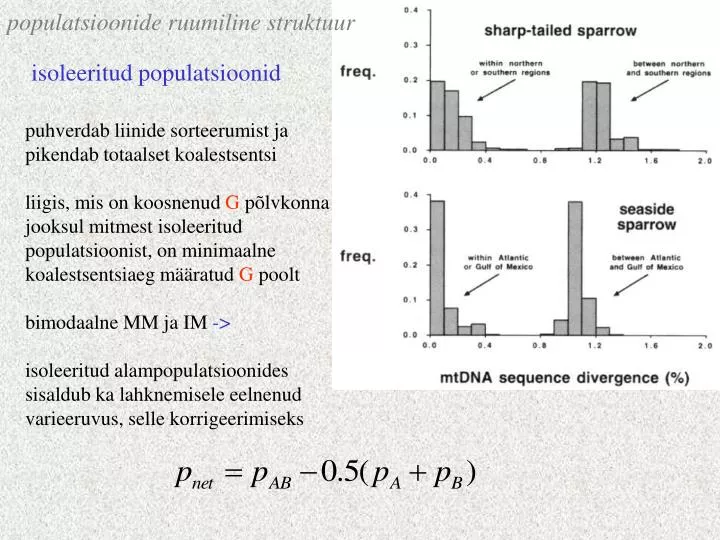
populatsioonide ruumiline struktuur. isoleeritud populatsioonid. puhverdab liinide sorteerumist ja pikendab totaalset koalestsentsi liigis, mis on koosnenud G põlvkonna jooksul mitmest isoleeritud populatsioonist, on minimaalne koalestsentsiaeg määratud G poolt

E N D
populatsioonide ruumiline struktuur isoleeritud populatsioonid puhverdab liinide sorteerumist ja pikendab totaalset koalestsentsi liigis, mis on koosnenud G põlvkonna jooksul mitmest isoleeritud populatsioonist, on minimaalne koalestsentsiaeg määratud G poolt bimodaalne MM ja IM -> isoleeritud alampopulatsioonides sisaldub ka lahknemisele eelnenud varieeruvus, selle korrigeerimiseks
min tAB max tAA ruumiline struktuur: genealoogiline lähenemine liinide sorteerumise tulemusena võimalikud kolme tüüpi topoloogiad: distantsarvutuslikult:
inimese polüfüleetiline mitokondripuu Euraasias
emaliinide sorteerumine põlvkondade lõikes polüfüülia parafüülia monofüülia G<NFe G>4NFe kehtib hästi segatud algpopulatsioonist juhuslikult võetud rajajate (founder) korral
vikaarsus ja eelnev genealoogiline ajalugu lahknemine on kooskõlas eelneva ajalooga eelnev ajalugu ei vasta fülogeograafilisele lahknemisele perifeerne lahknemine, millega A saab B suhtes parafüleetiliseks
parafüleetilisuse tagamaad ebatäielik liinide sorteerumine "gene-flow-early" parafüleetilised harud sama pikad kui monofüleetilised sekundaarne geenimigratsioon "gene-flow-late" äratuntav eelmisest kui migratsioon on suhteliselt hiljutine: parafüleetilised harud on lühikesed
menhaden fish phylogeography kaks morfoloogiliselt eristuvat liiki: Brevoortia tyrannusAtlanti rannik , Brevoortia patronus Mehhiko laht Atlanti ranniku kalad parafüleetilised Mehhiko lahe omade suhtes ebatäielik sorteerumine mõned Atlandi isendid identsed lahe isenditega hiljutine migratsioon lahest rannikule
lühiajaline isolatsioon ja rekoloniseerimine - vastupidi, vähendavad mitmekesisust ja totaalset koalestsentsiaega sisuliselt peegeldab mitmekesisus tuum- populatsiooni genealoogilist varieeruvust vaatamata geograafilise leviala ja summarse effektiivse populatsiooni suurusele. kehtib reegel, et kui lokaalsete populatsioonide väljasuremiseks kuluv aeg generatsioonides on väiksem kui vabalt ristuvate täiskasvanute arv, siis väljasuremise ja rekoloniseerumise vaheldumine inhibeerib lokaalsete populatsioonide differentseerumist triivi läbi seda effekti mõjutab ka populatsioonide vaheline migratsioon
population substructure Migration pt - frequency of allele after t generations p0 - initial frequency p - probability of the allele to be migrant = frequency in donor population m - migration rate per generation
Populatsion substructure MN Blood Group in Humans MN blood group system is one locus 2 alleles Frequencies have been measured in many human populations f(M) is 0.507 in white americans f(M) is 0.484 in black americans f(M) is 0.474 in west africans If the black population has been in America for 200-300 years, t = 10 generations and pt = p + (p0 - p)(1-m)t 0.484 = 0.507 + (0.474 - 0.507) (1-m)10 M = 0.035 or 3.5% of the genes at the MN locus in the ancestry of the black population have immigrated from the white population per generation In only 10 generations only one-third of the gene frequency difference has been removed by migration Migration is a much more powerful force in changing allele frequencies than is mutation
migratsioon island model (Wright 1931) - liik on jagatud võrdse suurusega populatsioonideks, mille vaheline migratsioon on igas suunas võrdse tõenäosusega stepping-stone model (Kimura 1953) - migratsioon leiab aset kõrvuti paiknevate populatsioonide vahel Nm, kus m on migratsiooni kiirus kui Nm on >1, siis migratsiooni ühtlustav mõju ületab triivi mitmekesistava toime 1) saare mudeli korral (Wright 1951) Fst alusel Nm(1-Fst)/4Fst 1 heterozygosity fixation index
migratsioon 2) populatsiooni-spetsiifiliste alleelide (private alleles) kaudu Nm rehkendus (Slatkin 1985) sõltuvad ajaloo sügavusest
migratsioon 3) kladistiline meetod (Slatkin and Maddison 1989) - säästu kriteerium hindamaks väikesimat migratsiooni sündmuste arvu (s) vastavalt puu topoloogiale puudused: 1) eeldus, et populatsioon on saavutanud triivi ja migratsiooni tasakaalu. See aga võib olla pikk, kui Ne on suur ja m väike: G=1/(2m+1)/2Ne)) 2) klassikalised hinnangud kehtivad liigi keskmisele, mitte vastavate, kontaktis olnud populatsioonidele 3) vastavalt graafikule, on raske kvantiteerida Nm; näiteks Fst=0.1 korral, kas tuhanded või mõned üksikud migrandid
migratsioon puudused: 4) Nm sõltuvalt populatsiooni suurusest mõjutab demograafiat väga erinevalt teised distantsarvutused: a) autocorrelation analysis (Sokal, Slatkin) b) PC analysis (Cavalli-Sforza) c) multidimensional scaling (Lessa) triivil põhiline kaal distantsmeetodites ei kasuta kogu informatsiooni, mis on arvesse võetav DNA järjestuste puhul
fülogeograafilised statistikud liigisiseste DNA järjestuste puhul puu konstrueerimine - MP eelsitatud kuna pole nii suurt kaalu pikkadel harudel ja kiiruste varieeruvusel kui liikide vahelistes suhetes Neigeli lähenemine (Neigel 1991; Neigel and Avise 1993) liigisiseste kiiresti evolutsioneeruvate molekulide korral (nagu mtDNA) pole tasakaaluline olek triivi ja migratsiooni vahel tõenäoline. Neigel käsitleb seega mittetasakaalulist populatsiooni kauguselise isolatsiooni (isolation by distance) mudeli korral. eeldatavalt eksisteerib korrelatsioon liini vanuse ja tema geograafilise leviku vahel. Erinevalt spetsiifiliste alleelide meetodist ei eeldata seost alleelisageduste ja nende divergeerumise aja vahel ega piirduta tähelepanuga haruldastele alleelidele. Meetod uurib puu kõigi harude ja haplotüüpide geograafilist levikut sõltumata hargnemiste sügavusest.
neigeli meetod liinid levivad (disperse) ruumis G põlvkonna jooksul samm haaval tütred levivad emadest ühe põlvkonna jooksul F kaugusele varieeruvusega F2 kui ühe ja sama haru kaks tippu koalestseeruvad G põlvkonda tagasi, siis nende vahele kuhjunud ruumilise kauguse varieeruvus on määratud G2=2GF2 ja ühe põlvkonna kohta levimise kaugus väljendub kui F=(F2/2G)1/2
Neigel 1991 rehkendas mtDNA phylogeograafilise andmestiku alusel leviku kiiruseks 200 m põlvkonna kohta - langeb enam-vähem kokku kohapeal mõõdetud 250 m sünnipaik-paarituskoht kaugusega puudusteks: 1) eeldab juhuslikku valimit 2) eeldab kella 3) üldine, mitte spetsiifiline - vaatamata puu struktuurile 4) eeldab piiranguteta levikut (barjäärideta)
Nested clade analysis A. Templeton 1992, 1995, 2000 Crandall 1996 1) hierarhilise struktuuriga kladogramm, juurimata (minimum spanning tree) 2) Dc - clade distance klaadi geogr. leviku määr Dn - nested clade distance klaadis oleva indiviidide keskmine geograafiline kaugus hierarhiliselt kõrgema klaadi indiviididest (keskmiselt) 3) permutatsioonid statistikud http://bioag.byu.edu/zoology/crandall_lab/geodis.htm
nested clade analysis Dc - geograafiline raadius väikseimale ringile, mis haarab kõiki indiviide Dn - kaugus ühisest keskmest haplotüüp A geograafiline leviala haplotüüp B geograafiline leviala statistiline analüüs nende parameetrite võrdlemiseks GeoDis programmis
nested clade analysis järeldused.., tõlgendused..
näide Prochilodus kalade fülogeograafia Lõuna-Ameerikas Andide kerkimine umbes 10 MYA http://golab.unl.edu/publications/Sivasundar2001/Sivasundar2001.html
mtDNA ATPaasi 6nda ja 8nda subühiku (840 bp) ML puu CR ML puu
klaadidel 2-3, 2-6, 3-2 puudub statsitiliselt oluline seos geograafilise leviku ja genealoogia vahel, ülejäänutel seos oluline, mis viitab levikulisele ekspansioonile (range expansion); klaad 2-1 Parana jõgikonnas 2000 km levialaga - seal puudub populatsiooni sisene struktuur
tagasimigratsioon Founder analysis doonorpopulatsioon retseptorpopulatsioon
1. ebatäielik valim 2. liinide sorteerumine Founder analysis founderite (rajajate) tuvastamine 1. kattuvad haplotüübid (match) 5. vahefaunder retseptoris 2. vahefaunderi kadu retseptoris ja kattuva tüübi kadu 3. kadunud faunder mõlemast 6. homoplaasia 4. kadunud faunder retseptoris
Founder analysis Forster et al. 1996 Ameerika indiaanlaste rajajaliinide tuvastamine tingimused: 1) ühised liinid Ameeriklaste ja Aasialaste vahel 2) rajajaliinid peaksid olema puu kesksetes sõlmedes, mitte perifeerias (paralleelmutatsioonid) 3) rajajaliinid peaksid olema laialdase geograafilise levikuga ning neist tulenevad harud kitsama levikuga -------------------------------------------------------------------------------------------------- Analüüs peaks tabama enamuse suurimatest (sorteerumine ja valim) rajajatest eeldustel, et a) migratsioon on mutatsioonikiiruse suhtes võrdlemisi hiljutine b) retseptorpopulatsioon on ekspandeerudes säilitanud enamuse liinidest
Founder analysis 1) ühised liinid: i) eralda kokkulangevad liinid kui faunderid ii) neist 1-sammu kaugusel olijad ei ole faunderid iii) faunderitest 2-e ja enama sammu kaugusel olevad liinid võivad omada valimis puuduvat vahefaunderit b) c) a)
Founder analysis 2) kesksed sõlmed: i) topiary pruning (Wills 1996) - stepwise mismatch analysis 1st step: in all 1-step differences replace character states with majority consensus
pärast pügamist 8nda tasemeni topiary pruning NJ puu enne pügamist
Founder analysis 2) kesksed sõlmed: i) topiary pruning (Wills 1996) probeem: pöördmutatsioonid ja majority consensus
Founder analysis kliki konsensus 2-sammuline klikk 4-sammuline klikk ii) median pruning (star contraction; Forster et al. unpublished) lokaalsed konsensused e. kliki konsensused klikk sisaldab endas kõiki maksimaalselt n sammu kaugusel olevaid liine
Founder analysis Forster et al. 1996 11 ühist, kattuvat liini 309 Aasia ja 472 Ameerika indiaanlase mtDNA HVS-1 ühises andmestikus + kaks keskset alamfaunderit Ameerikas (C, D1)
Founder analysis Forster et al. 1996 haplogrupp A2 Ameerikas mitmed alamfaunderid, seejärel enamvähem tähesarnane
Founder analysis geograafiline alamstruktuur lähtuvalt põhifaunderitest
Founder analysis Forster et al. 1996 klastrite ekspansiooniajad geograafiliselt jagatud populatsioonides S>3/2 kalibreerimine: A2 klastri diversiteet seotud põhjas noorema drüüasega (11,300 aastat)
Founder analysis Sykes et al. 1995 Polüneesia asustamine Grupp I ja III lähtuvalt Kagu-Aasia saarestikust Grupp II lähtuvalt Melaneesiast
Founder analysis B M11 tõenäoliselt Polüneesias või teel sinna tekkinud mutatsioon - dateerimaks tüüpiline Uus-Guineas F mitte-sage klaster, ebapiisav valim
Founder analysis Sykes et al. 1995
Founder analysis Richards et al. 2000 f0 rakendatud kriteeriumid faunderanalüüsiks: f1 , f2 piirangud: fs fs'
Founder analysis Richards et al. 2000 näide erinevate piirangute mõjust faunderite defineerimisel klassifitseerub faunderina iga piirangu korral Lähis-Idas (LI) puuduvad alamharud, ei klassifitseeru LI-st tuleneva faunderina
Founder analysis Richards et al. 2000 viis eeldust: 1) iga klaster on seotud ühe kindla migratsioonifaasiga 2) iga klaster ekspandeerus Euroopas kohe pärast rajaja saabumist (tähekujuline puu, mille sügavus peegeldab migratsiooni toimumise aega) 3) mutatsioonikiiruse hinnangud on täpsed 4) kõik mutatsioonid on fülogeneetilise analüüsiga topoloogiliselt lahendatud (reaalselt paralleel- ja tagasimutatsioonid) 5) faunderite järjestused on õieti tabatud
Founder analysis Richards et al. 2000 Haplogruppide jaotumine vastavalt migratsioonifaasidele
Founder analysis Richards et al. 2000 Üldsageduste jaotumine vastavalt piirangutele erinevus: tagasimigratsioon, paralleelmutatsioonid
Eksami küsimused !!! http://www.evolutsioon.ebc.ee/molevol/






















