Download

1 / 47
470 likes | 758 Views
Inhibition of Ethanol Neurotoxicity by Treatment with Growth Factors and Estrogen.

E N D
Inhibition of Ethanol Neurotoxicity by Treatment with Growth Factors and Estrogen Jason A. Zell, M.S., Jeremy R. Montague, Ph.D., Tomas F. Lopez, B.S., and Laura M. Mudd, Ph.D. 1999. Inhibition of Ethanol Neurotoxicity by Treatment with Growth Factors and Estrogen. McGill Journal of Medicine 5:13-24. By: Dawn Barclay
Purpose • To determine if Nerve Growth Factor (NGF), Basic Fibroblast Growth Factor (bFGF), Insulin-like Growth Factor (IGF-I), Insulin-like Growth Factor (IGF-II), or Estrogen (Es) could significantly inhibit the detrimental effect of ethanol on survival of cortical neurons and the concentration necessary for a significant effect.
Background • What is Fetal Alcohol Syndrome (FAS)? • Why the rat as a model system? • How does the rat compare with the human developmentally? • Why cortical neurons? • Why a bilaminar culture technique? • Why study NGF, bFGF, IGF-I, IGF-II, or Es?
What is FAS? • Intrauterine exposure of the human fetus to ethanol causes a neurotoxic syndrome. • Decreases in mental capacity and delayed maturation in humans following fetal alcohol exposure are associated with abnormalities in the number and structure of neurons throughout the cortex and other brain areas (West, 1986).
Why the rat as a model system? • The rat is a mammal. • FAS has been studied extensively on this model system. • The rat is commonly used as a model system for human development.
How does the rat compare with the human developmentally? • Days 1-9 in the rat correspond to the 1st trimester of human development.* • Days 10-21 in the rat correspond to the 2nd trimester of human development.* • Postnatal period of days 1-10 corresponds to the 3rd trimester of human development.* *(West, 1987)
In Vitro vs. In Vivo • In vitro-within glass or in an artificial environment.* • In vivo-within the living body.* • The use of a cell culture technique namely bilaminar cell culture technique to avoid some of the complicating factors found in an in vivo study. *http://www.graylab.ac.uk/cgi-bin/omd?action=Home&query=
Why a bilaminar culture? • Maintains separation between neuronal and glial plane while still allowing interaction. • This interaction allowed the experiments to be performed in the absence of serum. • Ultimately, allowing the experimenters to more accurately quantify both the concentration of growth factors/estrogen and ethanol the neuronal plane was exposed.
Why cortical neurons? • Ethanol causes decreased neuronal survival in the rat central nervous system (CNS) (Miller, 1995). • Miller, 1990 has shown that there is a 33% decrease in cortical neurons in ethanol exposed rat pups versus control. • West, 1986 showed that there are hypertrophic effects correlated with ethanol exposure such as alterations in the size of cell soma or nuclei, as well as dendritic length, density and branching patterns of neurons.
Neuron *Lodish, 2000
Glia *Lodish, 2000
Nerve Growth Factor (NGF) • Neurotrophin-associated with the mammalian cerebral cortex (Allendoerfer, 1990). • NG F utilizes a signal transduction pathway to act upon its target cell, specifically a TrkA receptor (Klien, 1991). • Followed by phosphorylation of key substrates that modify the physiology of the cell (Maher, 1998).
NGF cont… • NGF has been linked to neuronal survival in the post mitotic CNS (Downen, 1993). • NGF has been shown to increase neurite outgrowth (Hundle, 1995).
Insulin-like Growth Factor I & II (IGF-I & IGF-II) • IGF-I and IGF-II are found in the brain (LeRoith, 1989). • IGF-I appears to be involved in myelination and brain growth in vivo (Carson, 1993). • Genetic alterations of IGF-I result in reduced brain size, decreased myelination in the CNS, and losses of particular subpopulations of neurons (Beck, 1995).
IGF-I & IGF-II cont… • Kar, 1993 showed an increase in both IGF-I and IGF-II receptor levels following damage to the entorhinal cortex suggesting involvement with neuronal repair. • After ethanol exposure, expression of IGF-I and IGF-II in the fetal rat brain decreases (Singh, 1996).
Basic Fibroblast Growth Factor (bFGF) • Peptide and receptor are found within the cortex (Tagashira, 1995 and Weise 1993). • Differentiative vs. Mitogenic (Weise, 1993). • Protective role in CNS, mRNA increased after cortical injury (Yuguchi, 1994). • Increase in bFGF increased resistance to hypoxia (Akaneya, 1994).
Estrogen (Es) • Estrogen and receptors are found within the cortical tissue (Simerly, 1990). • Gibbs, 1994 showed that estrogen treatment decreases the incident and slows onset of Alzheimer's disease. • Gender differences in the onset and severity of neurodegenerative diseases such as schizophrenia (Hafner, 1993). • Suggests Es may have a neuroprotective effect on neurons within the brain.
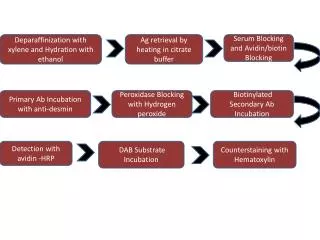
Preparation of Glial Cultures • Prepared from E16 Sprague Dawley rat cortices. • Glial dissection carried out 14 days prior to neuronal. • Plated in 60mm tissue culture dishes 6-8 days prior to neuronal dissection. • 40-60% confluence necessary for neuronal survival.
Preparation of Neuronal Plane • Cortical neurons were obtained from E16 Sprague-Dawley rat brains. • Neurons were plated on silane-coated glass coverslips with paraplast feet. • 3x104 cells/coverslip. • After 2 hours, coverslips were inverted over the 40-60% confluent glial plane.
Preparation of Neuronal cont… • The two cellular planes (bilaminar culture) were separated by a distance of approx. 1 mm. • After one day in vitro, cytosine arabinofuranoside was added to cultures to prevent glial proliferation. • Cell were maintained in a humidified 37oC, 95% air/5% CO2 incubator.
Experiment 1-Ethanol Evaporation Experiment • What is the rate of evaporation of ethanol within the cell cultures? • To ensure treatment patterns were synonymous with binge drinking/chronic treatment.
Methods-Ethanol Evaporation Experiment • 45mM concentration of ethanol was added to cell cultures on DIV4. • 100 mL samples were taken at 0, 2, 4, 6 hours. • These samples were analyzed using UV spectrophotometry for absorbance data. • Results were compared to Sigma Diagnostic ethanol kit #5133.
Methods cont… • Values were converted to amount of ethanol per volume media (mM concentrations) • Experiment was carried out in triplicate. • Mean values and Standard Error of the Mean were calculated.
Results • 0 hours-45mM + 2.3mM. • 2 hours-42mM + 0.8mM. • 4 hours-37.4mM + 1.4mM. • 6 hours-29.6mM + 0.8mM. A 34.3% decrease in ethanol concentration was observed in the media after 6 hours.
Experiment 2-Survival Assay • Does NGF, bFGF, IGF-1, IGF-2, or Es have a neuroprotective effect on survival? • If so, does it correlate with DIV 4 or DIV 6?
Methods-Survival Assay • Control/No EtOH—DMEM only. • Control/EtOH only—45mM EtOH • Dish 1/NGF and EtOH—EtOH and 100 ng/mL. • Dish 2/bFGF and EtOH—EtOH and 5ng/mL. • Dish 3/IGF-1 and EtOH—EtOH and 10ng/mL. • Dish 4/IGF-II and EtOH—EtOH and 10 ng/mL. • Dish 5/Es and EtOH—EtOH and 10nM Es. • Note-All treatments groups used 45mM Ethanol and were treated on DIV1 and DIV4.
Methods-cont… • Cell viability was determined using fluorescence microscopy and intravital dyes fluorescein diacetate and propidium iodide. • Cells were counted and expressed as percentage of the total.
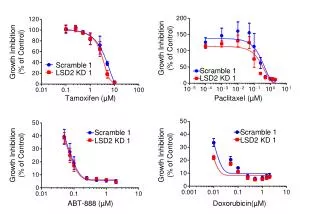
Results • Control/No EtOH—36.2+2.6% • Control/EtOH only—10.6+1.3% • NGF, bFGF, IGF-1, and Es showed a significant increase in cell survival over control/EtOH only. • Significance was assessed using a Dunnett’s test p<0.05.
Results • Control/No EtOH—22.7+2.3% • Control/EtOH only—3.7+1.3% • NGF, bFGF, IGF-1, IGF-II, and Es showed a significant increase in cell viability over control/EtOH only. • Significance was calculated using a Dunnett’s test p<0.05.
Experiment 3-Dose Dependence Experiment • What concentration of growth factor/estrogen facilitates the greatest neuroprotective effect? • Since physiological concentrations were used in survival assays, pharmacological concentration needed was the next logical question.
Methods-Dose Dependence Experiment • Three treatment groups per growth factor/estrogen were established. • Experiment were performed in triplicate. • Cells were treated DIV1 and DIV4. • Viability was assessed on DIV5. • Fluorescent microscopy was utilized. • FDA and PI were used as cellular markers, as described in survival assays.
Methods-cont… • Control/No EtOH • Control/EtOH only • NGF—100ng/mL, 10ng/mL, 1ng/mL & EtOH • bFGF—5ng/mL, 0.5ng/mL, 0.05 ng/mL & EtOH • IGF-I&II—10ng/mL, 1ng/mL, 0.1ng/mL & EtOH • Es—10nM, 1nM, 0.1nM & EtOH • Note: All growth factor/estrogen cultures were treated with 45mM EtOH.
Results • Control/EtOH only—4.6+0.6% • NGF—Maximal survival 100ng/mL-11.0+1.0% • Both 10ng/mL and 100ng/mL treatments were significant over control/EtOH only. • bFGF—Maximal survival 5ng/mL-17.7+4.3% • 5ng/mL treatment was the only significant treatment associated with bFGF.
Results-cont… • IGF-I—Maximal survival 1ng/mL-11.5+1.5% • Both 1ng/mL and 10ng/mL IGF-I treatment groups were significant over control/EtOH only. • IGF-II and Es did not produce significant results. • Significance was assessed using Dunnett’s test p<0.05.
Conclusions • The neurotoxic effect of chronic ethanol exposure can be attenuated by simultaneous treatment with NGF, bFGF, IGF-I, IGF-II, and Es. • On DIV4, survival was significantly increased over control/EtOH only in all groups except IGF-II. • Although by DIV6, all groups including IGF-II showed statistically significant increases in survival over control/EtOH only.
Conclusions-cont… • Logarithmic increases in growth factor concentrations were found to increase the inhibition of ethanol’s neurotoxicity at DIV5 in the NGF, bFGF, and IGF-I trials. • NGF did not show a linear increase in survival. • On the other hand, bFGF did show a dose dependent response. • IGF-I showed similar responses for two dosing regimes.
Discussion • Further exploration is needed to fully understand the mechanisms behind the increased survival conferred by the growth factor/estrogen treatments.
Questions? Comments?
References • Adamo M, Raizada MK, LeRoith D. 1989. Insulin and insulin-like growth factor receptors in the nervous system. Mol Neurobiol. 3(1-2):71-100. Review • Akaneya Y, Takahashi M, Hatanaka H 1994. Death of cultured postnatal rat CNS neurons by in vitro hypoxia with special reference to N-methyl-D-aspartate-related toxicity.Neurosci Res. 19(3):279-85. • Beck KD, Powell-Braxton L, Widmer HR, Valverde J, Hefti F. 1995. Igf1 gene disruption results in reduced brain size, CNS hypomyelination, and loss of hippocampal granule and striatal parvalbumin-containing neurons. Neuron 14(4):717-30. • Carson MJ, Behringer RR, Brinster RL, McMorris FA. 1993. Insulin-like growth factor I increases brain growth and central nervous system myelination in transgenic mice. Neuron 10(4):729-40.
References-cont… • Downen M, Mudd L, Roback JD, Palfrey HC, Wainer BH. 1993. Early nerve growth factor-induced events in developing rat septal neurons. Brain Res Dev Brain Res. 74(1):1-13. • Hafner H, an der Heiden W, Hambrecht M, Riecher-Rossler A, Maurer K, Loffler W, Fatkenheuer B. 1993 [A chapter in systematic schizophrenia research--the search for causal explanations for sex differences in age of onset]. Nervenarzt. 64(11):706-16. Review. • Hundle B, McMahon T, Dadgar J, Messing RO. 1995. Overexpression of epsilon-protein kinase C enhances nerve growth factor-induced phosphorylation of mitogen-activated protein kinases and neurite outgrowth. J Biol Chem. 270(50):30134-40. • Kar S, Baccichet A, Quirion R, Poirier J. 1993. Entorhinal cortex lesion induces differential responses in [125I]insulin-like growth factor I, [125I]insulin-like growth factor II and [125I]insulin receptor binding sites in the rat hippocampal formation. Neuroscience 55(1):69-80.
References-cont… • Klein R, Jing SQ, Nanduri V, O'Rourke E, Barbacid M. 1991. The trk proto-oncogene encodes a receptor for nerve growth factor. Cell 65(1):189-97. • Lodish H, Berk A, Zipusky SL, Matsudaira P, Baltimore D, Darnell JE. 2000. Nerve Cells. Molecular Cell Biology 4th edition. WH Freeman and Company, New York. • Miller MW. 1995. Effects of pre- or postnatal exposure to ethanolon the total number of neurons in the principal sersory nucleus of the trigeminal nerve: cell proliferation and neuronal death. Alcoholism clinical and Experimental Research 19:1359-1363. • Miller MW and Potempa G. 1990. Numbers of neurons in mature rat somaosersory cortex: effects of prenatal exposure to ethanol. Journal of Comparative Neurology 293:92-102.
References-cont… • Simerly RB, Chang C, Muramatsu M, Swanson LW. 1990. Distribution of androgen and estrogen receptor mRNA-containing cells in the rat brain: an in situ hybridization study. J Comp Neurol. 294(1):76-95. • Singh SP, Ehmann S, Snyder AK. 1996. Ethanol-induced changes in insulin-like growth factors and IGF gene expression in the fetal brain. Proc Soc Exp Biol Med. 212(4):349-54. • Tagashira S, Ozaki K, Ohta M, Itoh N. 1995. Localization of fibroblast growth factor-9 mRNA in the rat brain. Brain Res Mol Brain Res. 30(2):233-41. • Weise B, Janet T, Grothe C. 1993. Localization of bFGF and FGF-receptor in the developing nervous system of the embryonic and newborn rat. J Neurosci Res. 34(4):442-53.
References-cont… • West JR, Pierce DR. 1986. Perinatal alcohol exposure and neuronal damage. Pgs 120-157 in West JR Ed. Alcohol and Brain Development. Oxford University Press, New York. • Yuguchi T, Kohmura E, Yamada K, Wanaka A, Otsuki H, Sakaguchi T, Yamashita T, Tohyama M, Hayakawa T. 1994. Messenger RNA and protein expression of basic fibroblast growth factor receptor after cortical ablation. Brain Res Mol Brain Res. 25(1-2):50-6.