Download

1 / 33
330 likes | 406 Views
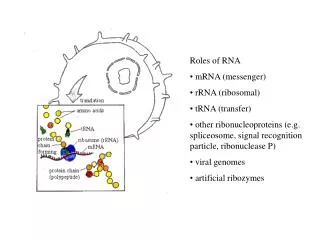
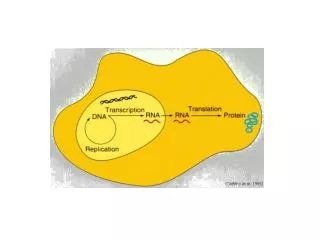
Structure and Folding of Ribosomal Frameshift-stimulating mRNA Pseudoknots David Giedroc Department of Biochemistry and Biophysics Texas A&M University. The Central Dogma…. DNA mRNA Protein. REGULATION: Transcriptional regulators; RNAi Metal Homeostasis/Resistance

E N D
Structure and Folding of Ribosomal Frameshift-stimulating mRNA Pseudoknots David Giedroc Department of Biochemistry and Biophysics Texas A&M University
The Central Dogma…. DNA mRNA Protein REGULATION: Transcriptional regulators; RNAi Metal Homeostasis/Resistance “A bioinorganic twist” transcription REGULATION: Ribosomal Recoding, e.g. -1 Ribosomal Frameshifting translation Our interests: Molecular Determinants of Biological Regulation
Ribosomal Recoding • Ribosomal or translational recoding: A programmed alteration in the usual triplet • decoding of the mRNA into protein by the elongating ribosome • Documented to occur in all organisms (bacteria to mammals to plants) • Translation recoding signals are embedded in the mRNA itself • Recoding comes in several flavors: • 1) Readthrough via stop codon (UAG) supression: fusion protein • 2) Bypass or hopping (T4 gene 60) • 3) Incorporation of nonstandard amino acids, including • selenocysteine (#21) and L-pyrrolysine (#22) via stop codon redefinition • 4) Frameshifting: change in reading frame to create a fusion protein • +1 PRF: XXX YYY ZZ XXX YYZ Z (antizyme: polyamine biosyn; E. coli RF2) • -1 PRF: X XXY YYZ XXX YYY Z (animal/plant RNA viruses; E. coli dnaX) From Baranov et al. (2002) Gene
Many RNA viruses employ -1 ribosomal frameshifting fs = 5% antiviral target protease P2 RdRP
[ [ [ [ Translational Recoding by -1 Frameshifting (e.g., PEMV-1) -1: 0: Frameshifting efficiencies typically range from 5-30% in RNA viruses
RNA Pseudoknot Folding Topology Example: Phage T2 gene 32 translational operator Du, Giedroc and Hoffman (1998) Biochemistry
Structural Diversity in RNA Pseudoknots Giedroc et al. (2000) J Mol Biol
L7/L12 Modeling the FS signal on the bacterial 70S ribosome (1JGO) Giedroc et al. (2000) J Mol Biol Plant et al. (2003) RNA
Overview of Translation eEF2 (EF-G)-catalyzed translocation perturbed by the downstream pseudoknot Pseudoknot kinetically enhances partitioning of the elongating ribosome into the new -1 reading frame duringtranslocation [Namy et al. (2006) Nature].
EM structure of a stalled 80S ribosome-pseudoknot complex mRNA channel
Hypothesis: Some general feature(s) of pseudoknot structure, stability and/or kinetic lability are major determinants for frameshift stimulation Our approach: Systematically investigate the structures, stability determinants of a group of closely related -1 PRF-stimulating pseudoknots Objective: Identify key molecular features that modulate frameshift stimulation, independent of folding Evaluate the functional importance of these interactions in detail in a suitable mechanistic assay
Plant Luteoviruses Mosaic yellow pattern on leafs Enations Symptoms of infection with pea enation mosaic virus (PEMV-1) Typical luteovirus particles Smith and Barker (1999) The Luteoviridae (CABI)
Proposed 2º structures of BWYV, PEMV-1 and ScYLV P1-P2 pseudoknots Beet Western Yellows Virus Pea Enation Mosaic Virus (RNA-1) Sugarcane Yellow Leaf Virus BWYV infection of escarole
1.6 Å structure of the BWYV pseudoknot S2 5’ L2 A21 C22 U13 (L3) A23 A24 S1 A25 C8 L1 3’ Su, Egli, Rich et al (1999) Nat Struct Biol
Sugarcane Yellow Leaf Virus P1-P2 RNA Pseudoknot as a Structural Target Slip-site
NMR Spectroscopy as a Biomolecular Structural Tool Texas A&M (500, 600 MHz:) The Scripps Research Institute (900 MHz) Large fixed Bo: Nuclear magnets (protons, etc.) align in the magnetic field, and absorb radiofrequency energy; the decay of this excited state is a strong function of structural environment of individual atoms.
The predicted ScYLV P1-P2 mRNA pseudoknot adopts a well-folded PK conformation H5(C5C4N)H: NH2 protons of C27 reside in an unusual environment Cornish, Hennig, Giedroc (2005) Proc Natl Acad Sci
Loop L2 adenosine amino protons are resolved and protected from solvent exchange Cornish, Hennig, Giedroc (2005) Proc Natl Acad Sci
Solution structure of the ScYLV P1-P2 RNA pseudoknot 50±14º Cornish, Hennig, Giedroc (2005) Proc Natl Acad Sci
Solution Structure of the ScYLV P1-P2 RNA Pseudoknot L2 5’ S1 A13 C25 S2 G9 (L1) 3’ Cornish, Hennig, Giedroc (2005) Proc Natl Acad Sci
L2 S1 A13 C8+ S2 C25 G9 (L1) Cornish, Hennig, Giedroc (2005) Proc Natl Acad Sci
ScYLV Pseudoknot: Five consecutive base triples cis-Watson-Crick/sugar edge base pairing Hoogsteen base pairing *See also HCV IRES (Kieft et al., 2002) *See also A riboswitch (Serganov et al., 2004); G riboswith (Batey et al., 2004) [C•(U-A)] Cornish, Hennig, Giedroc (2005) Proc Natl Acad Sci
CURVES analysis of BWYV, PEMV-1 and ScYLV pseudoknot topologies BWYV S1 • Helical over-rotation: 89º • Horiz. displacement: 5.5 Å S2 PEMV-1 A-form coaxial helices S1 • Helical over-rotation: 98º • Horiz. displacement: 5.0 Å S2 ScYLV S1 • Helical over-rotation: 103º • Horiz. displacement: 7.6 Å S2
-1 Frameshift Stimulation by the ScYLV P1-P2 RNA Pseudoknot The ≈2.5-fold difference in FS stimulation between ScYLV and BWYV pseudoknots derives entirely with a 3’ C A substitution in loop L2 Cornish, Hennig, Giedroc (2005) Proc Natl Acad Sci
The structures of the C27A and WT ScYLV pseudoknots are essentially identical… …despite easily measurable structural perturbations at the helical junction region Cornish, Stammler & Giedroc (2006) RNA
The C27A ScYLV pseudoknot is destabilized relative to the WT RNA Cornish, Hennig, Giedroc (2005) Proc Natl Acad Sci
Helical junction pairwise coupling free energies (d) of WT and C27A pseudoknots Cornish & Giedroc (2006) Biochemistry
The mechanical model for stimulation of -1 PRF during translocation Ian Brierley, Univ of London Namy et al. (2006) Nature Prediction: More stable pseudoknots would be more effective frameshift-stimulators, generally consistent with our findings. However,stabilizing interactions localized in the helical junction region appear far more important. Hypothesis: Helical junction interactions may function as GATEKEEPERS (kinetic barrier) to ribosome-mediated pseudoknot unwinding.
Another perspective on ribosome-mediated unfolding during -1 PRF
So the moral of the story is……. Embrace Physical Chemistry!! NIH Predoctoral Training Programs in Biophysical Chemistry, Chemistry-Biology Interface
Conclusions Plant viral -1 frameshift-stimulating mRNA pseudoknots adopt unique triple helical architectures characterized by numerous loop-stem (L1-S2 & L2-S1) base triple (quadruple) interactions. Despite significant differences in the helical junctions among all three luteoviral pseudoknots, their global folds are remarkably similar. A major determinant for modulating frameshifting efficiencies by luteoviral pseudoknots is the 3’ nucleotide in loop L2. We propose that the helical junction functions as a kinetic barrier to ribosome-mediated pseudoknot unfolding. Ground- state structure is a poor predictor of frameshift-stimulation. These variant pseudoknots will be excellent tools with which to mechanistically probe how pseudoknots stimulate frameshifting (laser-based optical tweezers).
Acknowledgments Dr. Carla Theimer Dr. Paul Nixon Dr. Peter Cornish Suzanne Stammler Lichun Li Saritha Suram Dr. Raza Khan Dr. Mirko Hennig The Scripps Research Institute Dr. David W. Hoffman University of Texas at Austin NIH NSF Texas Higher Education Coordinating Board Dr. Peter Cornish