Download

1 / 25
280 likes | 1.22k Views
Chromosomes. Human Genome Basics. Most human cells are diploid , having 2 copies of each chromosome, one from each parent. The gametes , sperm and egg, are haploid , with 1 copy of each chromosome.

E N D
Human Genome Basics • Most human cells are diploid, having 2 copies of each chromosome, one from each parent. • The gametes, sperm and egg, are haploid, with 1 copy of each chromosome. • Humans have a haploid number n of 23, meaning that there are 23 chromosomes in a complete haploid set. Thus, gametes have 23 chromosomes and diploid cells have 46. The pairs of chromosome in a diploid cell are called homologues. • The X and Y chromosomes are sex chromosomes, and the other 22 chromosomes are called autosomes. • Normal males are XY and normal females are XX. Thus, sperm cells either contain an X or a Y. Eggs always contain an X chromosome.
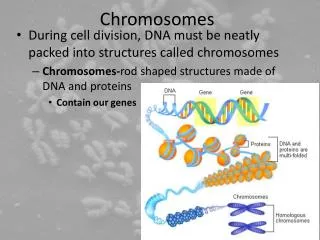
Cell Cycle and Chromosomes • A chromatid is a single DNA molecule along with supporting proteins and RNA molecules. • Chromosomes spend most of their time in the cell as single chromatids, the monad state. When a cell is getting ready to divide, the DNA in the chromosomes is replicated and the chromosomes become dyads, two chromatids held together at the centromere. • The dyad state is what is typically seen in pictures of chromosomes, but it only occurs briefly in the life of the cell. During most of interphase, chromosomes are in the monad state.
Cell Cycle • The cell cycle is a theoretical concept that defines the state of the cell relative to cell division. • The 4 stages are: G1, S, G2, and M. • M = mitosis, where the cell divides into 2 daughter cells. The chromosomes go from the dyad (2 chromatid) form to the monad (1 chromatid) form. That is, before mitosis there is 1 cell with dyad chromosomes, and after mitosis there are 2 cells with monad chromosomes in each. • S = DNA synthesis. Chromosomes go from monad to dyad. • G1 = “gap”. Nothing visible in the microscope, but this is where the cell spends most of its time, performing its tasks as a cell. Monad chromosomes • G2 (also “gap”). Dyad chromosomes, cell getting ready for mitosis. • G1, S, and G2 are collectively called “interphase”, the time between mitoses
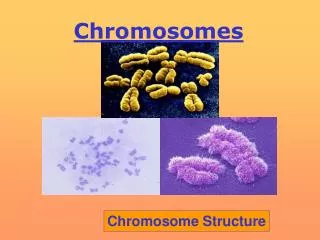
Chromosome Structure • DNA is long and thin and fragile: needs to be packaged to avoid breaking. • Lowest level is the nucleosome, 150 bp of DNA wrapped 1 3/4 times around a core of 8 histone proteins (small and very conserved in evolution). A string of beads. • The nucleosomes coil up into a 30 nm chromatin fiber. This level of packaging exists even during interphase. • During cell division, chromatin fibers are attached in loops of variable size to a protein scaffold. The DNA probably attaches at specific AT-rich areas called scaffold attachment regions. • The loops may be functional units: active vs. inactive in transcription. • Further coiling gives the compact structures we see in metaphase.
Centromeres • Sometimes called the “primary constriction” on a chromosome, based on microscopic appearance. • The centromere is the attachment point for the spindle. Acentric chromosomes, which don’t have a centromere, don’t attach to the spindle and don’t end up in either nucleus after mitosis. • The centromere is a region of DNA on the chromosome. During cell division, a large protein structure, the kinetochore, that attaches to the centromere DNA sequences. The spindle proteins then get attached to the kinetochore. • The centromere is many repeats of a about 170 bp element (very difficult to clone in humans but well known in yeast). Called alpha-satellite DNA. • Centromere regions also contain large amounts of repeated sequence DNA and transposable elements (more on this later).
Origins of Replication • DNA replication starts at specific spots on the chromosome, the origins of replication. • In yeast, these sequences are called autonomously replicating sequences (ARS) and are short well defined sequences. • In humans, it seems that origins of replication can vary a bit between cells and under different conditions. May be many of these on each chromosome.
Telomeres • Telomeres are the DNA sequences at the ends of chromosomes. Chromosomes that lose their telomeres often fuse with other chromosomes or become degraded. There are telomere-binding proteins that protect the chromosome ends. • Telomeres are also needed to ensure complete replication of the DNA: the end-replication problem • DNA polymerase must have a double stranded primer region with a free 3’ –OH to build on. The primer is made of RNA, synthesized by primase. At the 3’ end of the chromosome, the RNA primer gets degraded, leaving a single stranded region of DNA. • In the next round of replication, one DNA molecule will be shorter than the other. • Process repeats, gradually shortening the chromosomes. • Thought to be a cause of cell mortality.
More Telomeres • Chromosome shortening is prevented by telomerase, an RNA/protein hybrid enzyme. • Telomerase has a short RNA that is used as a template for a reverse transcriptase: binds to 3’ end of chromosome, then synthesizes DNA extension. This extension acts as a template for regular DNA polymerase, keeping chromosome length intact. • Telomere sequences are multiple repeats of a highly conserved 7 base sequence.
Euchromatin and Heterochromatin • Euchromatin is the location of active genes (although many genes in euchromatin are not active: depends on cell type). During interphase euchromatin is extended and spread out throughout the cell. • Heterochromatin is darkly staining, condensed, and late replicating. Genes in heterochromatin are usually inactive. • Some heterochromatin is constitutive : always heterochromatin: especially around centromeres. Composed mostly of repeat sequence DNA. • Other heterochromatin is facultative: can be heterochromatin or euchromatin: e.g. inactive X chromosome in females., the Barr body.
Mitosis • Mitosis is ordinary cell division among the cells of the body. During mitosis the chromosomes are divided evenly, so that each of the two daughter cells ends up with 1 copy of each chromosome. • Prophase: --chromosomes condense --nuclear envelope disappears --centrioles move to opposite ends of the cell --spindle forms • Metaphase: --chromosomes are lined up on cell equator, attached to the spindle at the centromeres • Anaphase: --centromeres divide. Now chromosomes are monads --the monad chromosomes are pulled to opposite poles by the spindle. • Telophase: --cytokinesis: cytoplasm divided into 2 separate cells --chromosomes de-condense --nuclear envelope re-forms --spindle vanishes
Meiosis • Meiosis is the special cell division that converts diploid body cells into the haploid gametes. Only occurs in specialized cells. • Takes 2 cell divisions, M1 and M2, with no DNA synthesis between. • In humans, start with 46 chromosomes (23 pairs) in dyad state. After M1, there are 2 cells with 23 dyad chromosomes each. After M2 there are 4 cells with 23 monad chromosomes each. • Prophase of M1 is very long, with a number of sub-stages. • Main event in prophase of M1 is “crossing over”, also called “recombination”. • In crossing over, homologous chromosomes pair up, and exchange segments by breaking and rejoining at identical locations. • Several crossovers per chromosome, with random positions. This is the basis for linkage mapping. • Chromosomes that don’t recombine seem to have a high rate of non-disjunction (chromosome goes to the wrong pole). Maybe homologues are held together until anaphase by recombination proteins.
More Meiosis • Metaphase of meiosis 1 is very different from metaphase in mitosis (or M2). • In metaphase of M1, pairs of homologous chromosomes line up together. In mitosis and M2, chromosomes line up as single individuals. • Anaphase of M1: the spindle pulls the two homologues to opposite poles. However, the centromeres don’t divide, and the chromosomes remain dyads. • Telophase of M1: cytoplasm divided into 2 cells, each of which has 1 haploid set of dyad chromosomes • Meiosis 2 is just like mitosis. • In prophase, the chromosomes condense and the spindle forms. • Metaphase of M2: dyad chromosomes line up singly on the cell equator. • Anaphase of M2: centromeres divide, chromosomes are now monads which get pulled to opposite poles. • Telophase: cytoplasm divided into 2 cells. • After M2: total of 4 cells from the original cell. Each contains one haploid set of monad chromosomes
Chromosomes in the Microscope • Studied in metaphase cells, usually white blood cells or skin cells. • Technique: arrest cell division at metaphase with colchicine or colcemid (blocks spindle microtubules). Then, hypotonic treatment swells them and spreads out the chromosomes. • Picture is a karyotype: chromosome pictures cut out and sorted by hand, or by computer. • Length varies: longest is chromosome 1, shortest is 21 (should be 22, but mistakes were made early on). • Centromere position: centromere index: length of short arm divided by total length. Used to define metacentric, sub-metacentric, acrocentric. (No human telocentrics) • Bands seen with different stains, especially Giemsa stain, which produces G bands. R bands are “reverse Giemsa”, the light bands seen with Giemsa stain. Bands seem to derive from large scale variations in GC content.
Nomenclature • Short arm is p (petite) and long arm is q. • cen is centromere, ter is terminus (telomere): pter and qter. • “proximal” means closer to the centromere, and “distal” means father away from the centromere • Regions divided at major bands: p1, p2, p3, etc. Then each region is divided into lesser bands; p11, p12, etc (p-one-one, not p-eleven). Even smaller bands too: p12.1, etc.
FISH • Fluorescence in situ hybridization. • Hybridize a DNA probe labeled with a fluorescent marker to chromosomes, then visualize in fluorescence microscope. • See location of the gene: often can see sister chromatids even. • Chromosome painting: use many probes from a single chromosome (there is lots of unique DNA on each chromosome). Good for seeing rearrangements. • Picture is translocational Down syndrome. Two copies
Chromosome Abnormalities • Three basic types: • polyploidy: having more than 2 sets of chromosomes. • aneuploidy: having an extra copy or a missing copy of a single chromosome. (equal numbers of all chromosomes is euploid). • mixoploidy: having cell lines with different chromosomal constitutions. • mosaics: derived from a single zygote. • chimeras: derived from the fusion of 2 different embryos
Polyploids • Triploidy generally results from dispermy: fertilization of the egg by 2 sperm simultaneously. Occasionally survive to birth, but die shortly after. • why is triploidy lethal, since all chromosomes are present in equal numbers (euploid)? May be due to X inactivation: only a single X is active in the cell, so there is an imbalance between gene products from the X and gene products from the autosomes. • Tetraploidy (4 sets of chromosomes). Very rare and always lethal. Usually due to failure of first mitotic division: chromosomes replicate and divide, but all end up in the same nucleus.
Aneuploidy • Two causes: • non-disjunction: paired chromosomes both go to the same pole in meiosis instead of to opposite poles. • anaphase lag: a chromosome moves to the pole so slowly that it doesn’t get incorporated into the nucleus as it forms in telophase. • Effects: fairly well tolerated for the sex chromosomes, but bad for autosomes. All autosomal monosomies and trisomies have been seen, but most are spontaneously aborted.
Sex Chromosome Aneuploidy • 45,X (XO: Turner). No ovaries = no puberty or secondary sexual characteristics (treat with hormone replacements). Folds of skin at neck, short stature, widely spaced nipples. Normal intelligence, but maybe spatial abilities impaired. • XXX, XXXX, etc. tall, some menstrual irregularities, sometimes slightly reduced intelligence, but not marked. Usually only found by chance. • XXY (Klinefelter). feminized male: may develop breasts. Limb elongation (can be prevented by testosterone injections). Slight retardation in some but not in all. Many only discovered by chance: infertility is a common symptom, e.g. • XYY. Tall, acne, slightly retarded. More in prison and mental institutions than expected by chance, but this may be a secondary effect only. • YY without an X: Embryonic lethal: too many genes on the X, but very few on the Y.
Autosomal Aneuploidy • Trisomy 21 (Down). epicanthal folds and flat face led to old name of "mongolism". Also, protruding tongue, loose joints, heart defects, suppressed immune system, increased risk of Alzheimer's. Large risk increased with maternal age. 5% of cases are translocational. Large variation in mental abilities. • Trisomy 18 (Edward). Mostly die before birth; few live past age 1. Major abnormalities of heart and liver. Odd way of making a fist: fifth finger and thumb folded over the middle three. mental development arrests at about 6 month stage. • Trisomy 13. (Patau). rarely survive till birth, almost none past age 1. Eye problems: small or absent; sometimes 1 eye hole in center of face, with nose above (a Cyclops). Also cleft palate. Mentally at 6 month level.
Chromosome Structure Changes • Caused by chromosome breaks. Enzymes join broken ends randomly. Two or more breaks often means the wrong ends are attached. • Sometimes this results in acentric or dicentric chromosomes (0 or 2 centromeres). Acentrics are lost in cell division; they don’t end up n the nucleus. Dicentrics are sometimes pulled apart or otherwise are unstable in mitosis. • Sometimes rejoining the wrong ends can result in a broken, non-functional gene at the breakpoint. • Or, two genes fused to give an abnormal phenotype. 11p13, results from the fusion of the ABL oncogene on chromosome 9 with the BCR gene on chromosome 22. • A balanced chromosome abnormality has all of the normal diploid number of genes: no gain or loss of genetic material. Such people are normal in phenotype if none of the above problems pertains. Many balanced abnormalities produce aneuploid gametes, leading to sterility or abnormal offspring. • Unbalanced chromosome changes have extra or missing genes. Many genes need to be present in exactly 2 copies, and having 1 or 3 copies leads to abnormality. These often add up to a syndrome, a group of symptoms having a common cause. For example WAGR syndrome (WILMS TUMOR--ANIRIDIA--GENITOURINARY ANOMALIES--MENTAL RETARDATION) is caused by a deletion of 11p13. Each element of the syndrome is caused by a separate gene.
More Structural Changes • Robertsonian translocation. Also called a centric fusion. • Chromosome breaks near the centromeres in the short arms of 2 acrocentric chromosomes gives one translocation chromosome with both long arms and one with both short arms. • The short arms of acrocentrics often have no vital genes and so can be lost. • balanced, but offspring are often aneuploid • cause of translocational Down syndrome, a t(14/21) –long arms of chromosomes 14 and 21. • isochromosomes. Both arms of a chromosome are identical. Caused by unusual crossover event between sister chromatids.
Chromosomes from the Wrong Parent • The DNA of some genes is modified by adding methyl groups to some C bases. This is called imprinting. For some genes imprinting is different for male and female gametes, and the gene from the father doesn’t work the same as the gene from the mother, at least in the embryo. • Uniparental diploids: both sets of chromosomes from the same parent. All are very non-vaible: • paternal uniparental diploid. Egg loses its nucleus, gets fertilized by an X-bearing sperm, with first mitosis resulting in one diploid nucleus. Result is a hydatidiform mole, has external membranes and structures of an embryo, but no actual embryo. Can become cancerous. • maternal uniparental diploids. Unfertilized egg gets activate. Results in an ovarian teratoma, a disorganized mass of tissues often including hair, bones and teeth, but no external embryonic membranes. • Uniparental disomy: a single chromosome with both copied form one parent. Results from a trisomic cell losing a chromosome and thus becoming viable. Best case is PRADER-WILLI SYNDROME/ANGELMAN SYNDROME, where inheriting 2 copies of the mother’s chromosome 15q gives Prader-Willi and 2 copies of the father’s 15q gives Angelman.