Download
1 / 31
350 likes | 805 Views
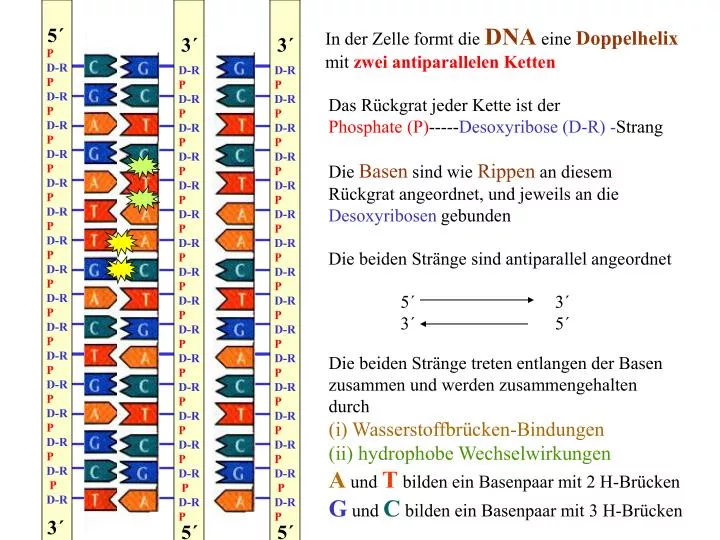
5´. 3´. 3´. P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R. D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P. D-R P D-R P D-R P

E N D
5´ 3´ 3´ P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P D-R P Das Rückgrat jeder Kette ist der Phosphate (P)-----Desoxyribose (D-R) -Strang Die Basen sind wie Rippen an diesem Rückgrat angeordnet, und jeweils an die Desoxyribosen gebunden Die beiden Stränge sind antiparallel angeordnet 5´ 3´ 3´ 5´ 3´ 5´ 5´ In der Zelle formt die DNA eine Doppelhelix mit zwei antiparallelen Ketten Die beiden Stränge treten entlangen der Basen zusammen und werden zusammengehalten durch (i) Wasserstoffbrücken-Bindungen (ii) hydrophobe Wechselwirkungen A und T bilden ein Basenpaar mit 2 H-Brücken G und C bilden ein Basenpaar mit 3 H-Brücken
Regeln bei der Watson-Crick-Basenpaarung (i) Wasserstoffbrücken-Bindungen (ii) hydrophobe Wechselwirkungen
Regeln bei der Watson-Crick-Basenpaarung (i) Wasserstoffbrücken-Bindungen
biologisch releveant 12 - 29 kJ/mol Regeln bei der Watson-Crick-Basenpaarung (i) Wasserstoffbrücken-Bindungen 0.27-0.31 nm
5´ 3´ H-Brücken 3´ 5´ Ausbildung der Watson::Crick Basenpaare innerhalb der DNA-Doppelhelix Regeln bei der Watson-Crick-Basenpaarung die beiden DNA-Stränge treten entlang der Basen zusammen und werden zusammen- gehalten durch (i) Wasserstoffbrücken-Bindungen (ii) hydrophobe Wechselwirkungen die Basen auf den gegenüberliegenden Strängen stehen sich exakt gegenüber A und T ein Basenpaar mit 2 H-Brücken G und C ein Basenpaar mit 3 H-Brücken die Basen sind planar gebaut > Stapelung der Basenpaare in der Doppelhelix (´base stacking´)
Fragen aus der schriftlichen Physikumsprüfung G = C A+G = T+C A = T
Strukturprinzip der Doppelhelix Die DNA-Doppelhelix das Desoxyribose-Phosphat- Rückgrat bildet eine Helix-Struktur aus die Basen sind zum Innern der Helix gekehrt und die Desoxyribose/Phosphat-Reste liegen an der Außenseite die Ringebenen der Basen stehen senkrecht zur Helixachse die Zucker stehen fast im rechten Winkel zur Base die genetische Information liegt in der Abfolge der Basenpaare TAGCGT Leiter-Modell Ribbon-Modell
Die Struktur der DNA-Doppelhelix Raumfüllendes Kalottenmodell 1. Strang (5´) 2. Strang (3´) große Furche kleine Furche DNA-bindende Proteine können spezifisch an die DNA binden. Dabei lagern sich die Proteine in die große bzw. kleine Furche ein und bekommen dadurch Zugang zu den Basen. Dadurch ist die Erkennung einer spezifischen Basenabfolge möglich (z. B. Transkriptionsfaktoren, Restriktionsenzyme) rote Linien verbinden die Phosphat-Reste
GCGCGCGC Abfolge Zick-Zack Verlauf des Phosphodiester-Bandes Linksläufige Doppelhelix Rechtsläufige Doppelhelix Entspricht 99% der zellulären DNA Entsteht bei drastischer Abnahme des Wassergehalts Rechtsläufige Doppelhelix Basenpaare nicht senkrecht wie bei B-Form, sondern um 70° gekippt Offener Raum im Molekülinneren
Die Verdopplung der DNA 1953 haben Watson & Crick die drei-dimensionale Struktur der DNA -Doppelhelix aufgeklärt und sofort den Mechanismus der DNA-Replikation vorhergesagt
Die DNA-Replikation wichtig ist, daß die genetische Information (= Abfolge der Basensequenz in der DNA-Kette) während der Zellteilung exakt von der Mutterzelle auf die Tochterzelle übertragen wird Fehler können sich kastastrophal auf die Lebensfähigkeit der Zelle auswirken, oder Krebs entstehen lassen bei der Verdopplung fungieren beide Elternstränge der doppelsträngigen DNA als Matritze
bei der Verdopplung fungieren beide Elternstränge der doppelsträngigen DNA als Matritze
Identische Verdopplung Reißverschlußartige Trennung der beiden Elternstränge entlang der Basenpaare = Brechen der H-Brücken Semi-Konservative DNA-Replikation
A1 A1 A2 A2 Parentale DNA- Doppelhelix A1 A2 N1 N2 N1 A1 A2 N2 DNA-Moleküle nach einer Runde der Replikation Konservative Replikation Semi-Konservative Replikation Semi-konservative DNA-Verdopplung wurde durch Watson & Crick (1953) vorhergesagt, aber erst durch ein klassisches Experiment (1959) bewiesen von Meselson & Stahl
Der semi-konservativeReplikationsmechanismus wurde zunächst für E. coli gezeigt, und später für alle Organismen einschließlich Homo sapiens bewiesen
Verfüttern von 15N-Stickstoff Nährmedium an E. coli „Leichte DNA“ „Schwere DNA“ E. coli DNA enthält normalen = leichten 14N-Stickstoff DNA enthält jetzt ausschließlich schweren 15N-Stickstoff Rückführung der Bakterien in 14N-Stickstoff-Nährmedium für eine Verdopplungszeit von 20 min Anschließende Analyse der DNA ob schwer, leicht oder Hybrid durch CsCl-Dichtegradientzentrifugation
Mögliche Ergebnisse Experimentelle Befunde (Meselson & Stahl, 1958) konservativ Semi-konservativ S-S S-S S-S L-L S- L S- L Analyse der DNA ob schwer, leicht oder Hybrid durch CsCl-Dichtegradientzentrifugation S-S L-L L-L L-L S- L L-L S- L L-L Wachstum in 15N-Medium Schwere DNA (schwer = S) S-S 1. Verdopplung mit 14N-Stickstoff (leicht = L) S-L L-L S-L 2. Verdopplung mit 14N-Stickstoff (leicht = L)
e- Objektträger Das Beweis-Experiment, daß auch eukaryontische DNA semi-konservativ repliziert wird Dazu wurden Wurzelzellen mit [3H]-Thymidin (radioaktiver Wasserstoff!) durchmarkiert, isoliert und anschließend autoradiographisch untersucht Röntgen-Film mit AgBr-Emulsion Radioaktiv-markiert Probe (z. B.[3H]-Thymidin-markiertes Chromosom) beimb-Zerfall des Tritiums werden Elektronen frei, die den Röntgenfilm an den Stellen schwärzen (Ag-Körner), an denen die Chromosomen auf dem Film aufliegen. Genauigkeit der Lokalisierung: ca. 0.5 - 1.0 mm
A1 N1 A2 N2 DNA-Replikation in nicht-radioaktivem Medium Teilung DNA-Synthese in radioaktivem Medium [3H]-Thymidin 2n Autoradiographie-Bild Autoradiographie-Bild beide Schwester- Chromatide sind markiert nur ein Schwester- Chromatid ist markiert …….semi-konservative Replikation von eukaryontischer DNA in Wurzelzellen A1 A2 1n 2n
Die DNA-Replikation erfolgt bi-direktional Replikationsgabel • Die Replikation der DNA erfolgt nicht spontan, sondern wird durch Enzyme (DNA-Polymerasen) katalysiert • Grundsätzlich gilt, daß die DNA-Polymerasen die Replikation nicht an jeder beliebigen Stelle der DNA initiieren, sondern nur am Replikations-Startpunkt = “Origin“
linke Wachstumsgabel rechte Wachstumsgabel Ende der Replikation • am häufigsten findet man, daß die Replikation am Origin beginnt und sich von dort bi-direktional forstsetzt • dadurch entstehen2 Replikationsgabeln Wachstum in beide Richtungen • bei ringförmiger DNA (z. B. E. coli) genügt meist einOrigin. Die beiden dort entstehenden Replikationsgabeln verschmelzen schließlich auf der gegenüberliegendenen Seite des Origins nach erfolgter Replikation
die langen, linearen eukaryontischen Chromosomen (=lineare DNA) haben viele DNA-Replikations-Startpunkte (vermutlich einige tausende bis zehntausende Origins beim Menschen) • die bi-direktionalen Replikationsgabeln bewegen sich aufeinander zu, solange bis sich benachbarte Startpunkte begegnen • durch das Vorhandensein von vielenReplikons (Replikations-Startstellen) schafft es die eukaryontische Zelle die ca. 1000-fach größere DNA (vgl. E. coli > Mensch) in der erforderlichen Zeit zu replizieren
e- Autoradiogramm Ori 1 Ori 2 hohe niedrige hohe Radioaktivität hohe niedrigehohe Radioaktivität [3H]-markierte DNA • Der Nachweis der bi-direktionalen DNA-Replikation wurde durch Autoradiographie von radioaktiv markierter DNA, die aus Säugerzellen isoliert wurde, erbracht. • Markierung der DNA erfolgte mit [3H]-Thymidin, zunächst von niedriger Radioaktivität.Nach Beginn der Replikation wurde die Menge an radioaktivem [3H]-Thymidinerhöht. Anschließend wurden die Chromsomen autoradiographiert.
Sichtbarmachen von Replikationsblasen 1n DNA Replikationsblase 2n DNA Replikationsgabel • Elektronenmikroskopische Aufnahmen von sich replizierender DNA aus Drosophila-Zellen, die sich rasch teilen (= aktive DNA-Replikation). Die Pfeile zeigen Replikationsblasen, in denen bi-direktionale Replikation stattfindet.
OriC t1 t2 t3 Wanderungs-Geschwindigkeit der Replikationsgabeln • Zellen werden unterschiedlich lang mit [3H]-Thymidin markiert, bevor die DNA isoliert und autoradiographisch analysiert wird • durch die Länge der Schwärzung auf dem Röntgenfilm kann die Geschwindigkeit, mit der sich die Replikationsgabel entlang der DNA bewegt, bestimmt werden hohe niedrigehohe Radioaktivität Längeder radioaktiv markierten DNA(mm) in Abhängigkeit derReplikationsdauer
Kleine Rechnung bei E. coli dauert die Replikation 20 min (Genomgröße = 3 x 106 BP) bei 2 Replikationsgabeln mit einer V = 1000 BP/sec stimmen theoretischer und errechneter Wert überein 1200 sec x 1000 BP = 1.2 x 106 BP bei 2 Gabeln: 2.4 x 106 BP • Homo sapiens~ 100 BP/sec/Gabel • (Autoradiographie) • bei 3 x 109 BP/Genom benötigt man ca. 1000 Replikationsgabeln für die Replikation des gesamten Genoms (in ca.10 hr) Wanderungs-Geschwindigkeit der Replikationsgabeln Länge der DNA auf dem Autoradiogramm proportional zur Anzahl der replizierten Basenpaare > Bestimmung der Replikationsgeschwindigkeit = 1000 BP/sec/Gabel bei E.coli
Startkomplex - Primase - DNA-Polymerase Replikation Wie funktioniert der einzige “Origin of Replication“ in E. coli? OriC DnaA (ATPase) 30°C DnaB = Helikase DnaC ATP Erkennung Offener Komplex leichtes „Schmelzen“ der DNA „Prä-Priming“ Komplex