Download

1 / 28
280 likes | 437 Views
Ribosomes , translation in laser trap Following translation by single ribosomes one codon at a time. Wen et al Nature 452 , 598 (2008) Note Nobel this year for ribosome structure importance of structural work cited in intro to this paper!.

E N D
Ribosomes, translation in laser trap Following translation by single ribosomes one codon at a time. Wen et al Nature 452, 598 (2008) Note Nobel this year for ribosome structure importance of structural work cited in intro to this paper!
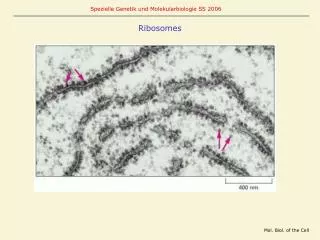
Translation performed by “ribosomes” ~ 20 nm 30S subunit 1540nt rna 21 proteins 50S subunit 120nt rna 2900nt rna 34 proteins
If you could watch this step by step, what might be the slowest (“rate- limiting”) steps? How many would there be? How would the ribosome move in time?
Experimental system mRNA made in vitro by T7 RNA pol from DNA templates designed to produce different “hairpins” Stall ribosome by withholding particular tRNA; then add it and “start the clock” d = F/k x record F vs x as function of t
Mixed aa’s Mostly V, E Alt. runs of V & E
Evidence that ribosome can read through ds stem (Fig. S3) 1,ribosome stalled at base of stem (F11); it takes 23pN to melt stem ( ) 2a, same as 1 but pull only to 20pN then, 2b add missing aa-tRNA, stem gradually melts due to ribosome reading through stem, melting it as it goes After some time, increase force: 2c shows no sudden increase in extension, i.e. no evidence of residual stem
Main result – at constant force, extension proceeds in steps – are steps convincing? Histogram of distances between all pairs of data points shows steps clearly; steps ~2.7nm (precision!) WLC model predicts 2.7nm @20pN <->6 bases =>melting 3 bases at a time Could you see 1.35nm step?
Expanded time course of step “Translocation” really = unwinding; translocation could occur during subsequent pause “The most parsimonious interpretation … is that unwinding and translocation … occur simultaneously…” What (besides step height) can one analyze in such traces?
Assume rate limiting events occur at random at rate k (per sec) p(1st event at time t)dt = k e-ktdt p-> k as t-> 0 Suppose 2 random, sequential events required, rates k1, k2 p(2nd event at time t)dt = p1st(t’)dt’ p2nd(t-t’) dt integrated over t’ from 0 to t. Find p(t) = [k1k2/(k2-k1)](e-k1t – e-k2t)dt For k1=k2, p = k2te-kt note p-> 0 as t-> 0 “Translocation” time dn best fit by 3 sequential events Does this identify what the events are? Bp melting?
With longer hairpin, pause times (extension rates) are quite variable different colors = identical RNAs where are pauses occuring? why?
Looking for correlations between sequence and pause sites Some clustering of pause sites (blue) downstream of “SD” AGGAGG. Complementary seq exists in 16S ribosomal RNA, previously implicated in ribosome binding to mRNA. Pauses eliminated by mutating to AAG, but this would also change binding to tRNA; is the D pause rate statistically significant?
Pauses can be terminated by pulling on the rna (promotes un- winding)
Dwell times also not fit by single exponential but this data less reliable: only first bin has too few occurrences, and only shortest dwell times were analyzed – long dwell times were attributed to unknown “other” phenomena.
Conclusions Single ribosome study allows detection of steps & pauses that can’t be studied in ensemble (bulk) studies because you can’t synchronize multiple ribosomes Not clear if steps are due to ribosomal translocation or ribosomal melting of 2ary structure in template If shorter distances can be reliably resolved (~ 1nm!), would be better to observe ribosome on less structured RNA template
Questions … extension Earlier Bustamante paper current paper Why does 5pN pull to ~ 90% of L on left but <10% on right?
Fig S1a – said to show that translation melts stem since stalling at sites further along stem results in smaller extensions at 23pN. Why are curves sequentially dis- placed to right? Rips at lower F (arrow heads) said to be due to 2ary structure 5’ of ribosome. Why are these variable?
Fig S1B. Pulling curves after various times of translation. Why does rip size decrease? Why do pre-rip portions of curves differ? What would you predict if structures were relaxed and re-pulled after ribosome had read through?
Fig. S4 VE274hp a:1, stall at 1st E, pull and relax; 2a pull to 17pn, hold at constant position, 2b add E-tRNA; 2c pull again. Why does F decrease (2b)? Why does extension increase as F decreases (panel b)?
Appreciate precision – detecting ~nm displacements in liquid, despite Brownian motion (damped to some extent by tightness of laser trap spring) They see 3 base step, hint of 1 base substeps Extract interesting observations even from “artifacts” e.g. peculiarly long pauses ? Shine Delgarnoseq related pauses, perhaps accidentally found because E encoded by GAN ? “clogging” of protein exit site in ribsosome by hydrophobic poly Val
Preview of next week’s topic Microtubules (from Howard & Hyman, Nature 422:753 (03) Part of cell “cytoskeleton” fibers often radiate out from site near nucleus called centrosome cell presumably stained w/ fluorescent anti-tubulin
Microtubules • Made of 2 proteins , tubulina and b, each ~ 500 aa • monomers -> a-bdimer (8nm long), • dimers -> ab-ab-ab... protofilaments • protofilaments -> tubes (microtubule) • ~13 protofilaments/tube circumference • tube diameter ~25nm, length up to 25 mm (1000:1!) • persistence length ~250mm; like Reif DNA tubes! • Tube is polarized: a at one end b at the other • grow in direction of + end
Tube growth and disassembly controlled by GTP binding btubulin can bind GTP and “hydrolyze” it to GDP + P In GTP-bound state, conform. favors dimer binding to + end This increases rate of GTP hydrolysis in adjacent dimer Internal dimers in GDP state prefer curved conformation but are held straight by binding to adjacent dimers in tube If end units hydrolyze GTP -> end dimers rapidly dissociate -> stochastic growth & retraction = dynamic instability
Label tubulin with green fluorescent protein in vivo -> Tubules continually grow, retract (“catastrophe”), regrow (“rescue”) “Dynamic instability” http://www.youtube.com/watch?v=ZL3_BwrB6AM http://www.youtube.com/watch?v=E1XczyCkN20&NR=1 GFP-label protein that binds microtubule +ends http://www.youtube.com/watch?v=9ICqcZ99qGk
Ends of tubules often associated with structures that get moved around inside cells by growth/retraction of tubules E.g. centrosome divides chromosomes duplicate + ends of tubules attach to chromosomes “catastrophe” pulls chromosome copies to what will become daughter cells
“Kinesin” proteins bind microtubules and “walk” along them, hydrolyzing ATP as they move Some kinesins have 2 heads, which alternately bind and release tubulin, “bipedal” walkers (recall DNA) What is role of ATP hydrolysis? http://www.youtube.com/watch?v=lLxlBB9ZBj4
If you immobilize kinesins on surface, add fluorescently labeled microtubules + ATP -> microtubules move along immobile “motors” http://www.youtube.com/watch?v=hqIRNekAdfA shows motor system only requires motor protein + ATP + track (tubulin)