Download

1 / 44
440 likes | 588 Views
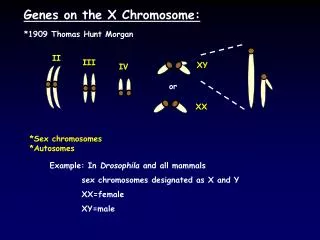
CHAP 3 POSITION DES GENES SUR LE CHROMOSOME ET LIAISON GENETIQUE. D’après « Génétique, les grands principes » Hartl et coll. Dunod 3 ème éd. 2003. Les gènes sont physiquement liés sur un chromosome Observation en conflit avec la 3 ème loi de Mendel???. Drosophile sauvage. [Oil blanc] w.

E N D
CHAP 3 POSITION DES GENES SUR LE CHROMOSOME ET LIAISON GENETIQUE
D’après « Génétique, les grands principes » Hartl et coll. Dunod 3ème éd. 2003 • Les gènes sont physiquement liés sur un chromosome Observation en conflit avec la 3ème loi de Mendel???
Drosophile sauvage [Oil blanc] w [Ailes atrophiées] m • m et w sont localisés sur le chromosome X. • Comment ségrègent les gènes correspondants l’un par rapport à l’autre ?
Ségrégation de gènes positionnés sur le chromosome X de drosophile P1 : ♀ [yeux blancs] x ♂ [ailes atrophiées] F1 ♀ [+] + ♂ [yeux blancs] ♀ [+] x ♂[yeux blancs, ailes atrophiées] Croisement test (test cross) Phénotype : Yeux blancs : 226 [w] Ailes atrophiées : 202 [m] Sauvages : 114 [+] Yeux blancs et ailes atrophiées : 102 [w,m] Total descendance : 644
Ségrégation de gènes positionnés sur le chromosome X de drosophile P1 : ♀ [yeux blancs] x ♂ [ailes atrophiées] w/w ; m+/m+ w+/- ; m/- F1 ♀ [+] + ♂ [yeux blancs] w/w+; m/m+ w/- ; m+/- ♀ [+] x ♂[yeux blancs, ailes atrophiées] w/w+; m/m+ w/- ; m/- Phénotype : Yeux blancs : 226 [w] Ailes atrophiées : 202 [m] Sauvages : 114 [+] Yeux blancs et ailes atrophiées : 102 [w,m] Total descendance : 644
w w+ w+ w+ w+ m m m m w w w w m+ m+ m+ m+ m+ w w m m w w+ m m+ P1 ♀ [yeux blancs] x ♂ [ailes atrophiées] F1 Gamètes de génotype parentaux pour m et w : Gamètes parentaux Gamètes de génotype recombinés pour m et w : Gamètes recombinés Gamètes
w+ w+ w+ w+ m m m m w w w w m+ m+ m+ m+ w w w w w w m m m m m m w w w w+ w+ w+ m m m m+ m+ m+ F1 Recombinés pour w et m : Recombinaison intrachromosomique Parentaux pour w et m : [œil blanc] 226 [ailes atrophiées] 202 [ailes atrophiées et œil blanc] 102 [sauvage] 114 Les gamètes recombinés entre w et m sont moins fréquents que ceux parentaux.
Fréquence de recombinaison: Nombre de chromatides (gamètes) recombinés (pour les marqueurs considérés) Nombre de gamètes total X 100 644 gamètes sont d’origine maternelle 114 + 102 sont recombinés entre w et m Fréquence de recombinaison = (216/644)x100 = 33,85% • Dans 33,85% des méïoses de la mère, il y a un CO entre w et m.
w+ w/w ; m+/m+ x w+/- ; m/- P1 w+/w+ ; m+/m+ x w/- ; m/- m+ w w w w w+ w+ w+ m m m m+ m+ m+ m+ w w+ m m m+ m+ w+ w+ m m+ w+ w+ 1/ Incidence de la configuration des allèles sur le chromosome sur la fréquence de recombinaison ♀ [yeux blancs] x ♂ [ailes atrophiées] ♀ [+] x ♂ [ailes atrophiées, yeux blancs] Femelle F1 w/w+ ; m+/m w/w+ ; m+/m w et m sont en configuration trans… w et m sont en configuration cis… m m+ w w m m+ w w …Ils « s’attirent »» au moment de la méïose …Ils se « repoussent » au moment de la méïose
w et m en trans w et m en cis w+ m+ w m w m w w+ m m+ Ségrégation de w et m dans la descendance mâle : 100 113 [w] [w] [m] [m] 101 101 [+] [+] 57 197 [w,m] [w,m] 51 199 322 597
w et m en trans w et m en cis w+ m+ w m w m w w w+ w+ m m m+ m+ w+ m+ Ségrégation de w et m dans la descendance mâle : 100 113 [w] P [w] [m] [m] 101 P 101 [+] [+] 57 R 197 w [w,m] [w,m] R 51 199 m 322 597 Frec (w,m) = 100x(57+51)/322 = 33,54%
w et m en trans w et m en cis w+ m+ w w m m w m w w w+ w+ m m m+ m+ w w+ m+ m w+ m+ Ségrégation de w et m dans la descendance mâle : 100 113 R [w] P [w] [m] [m] 101 P R 101 [+] w+ [+] 57 R 197 P m+ w [w,m] [w,m] R 51 199 P m 322 597 Frec (w,m) = 100x(57+51)/322 = 33,54% Frec (w,m) = 100x(101+100)/597 = 33,67% • La configuration ci ou trans des allèles n’a pas d’influence significative
2/ Incidence du sexe sur le taux de recombinaison c+ p+ c p La fréquence de recombinaison est-elle différente selon qu’elle est déterminée chez un mâle ou chez une femelle? [curly,œil pourpre] [sauvage] P1 F1 c/c+ ; p/p+
Test-cross (croisement test) Femelle F1 [+] x ♂ [c,p] Mâle F1 [+] x ♀ [c,p] [c,p] : 394 [c] : 104 [c,p] : 498 [p] : 106 [+] : 396 [+] : 502
c+ c+ c+ c+ p+ p+ p+ p+ c c c c p p p p c c c c c c c c c c p p p p p p p p p p c p+ c+ p Mâle F1 [+] x ♀ [c,p] Femelle F1 [+] x ♂ [c,p] c/c+ ; p/p+ c/c ; p/p c/c ; p/p c/c+ ; p/p+ 396 498 394 502 104 106 Frec(c,p) = 0 % Frec(c,p) = 21%
Le « sens » du croisement est donc particulièrement important chez la drosophile, car le mâle de la drosophile ne fait pas de recombinaison intra-chromosomique (pas de CO). • Chez les autres eucaryotes, il y a toujours une petite différence dans le taux de recombinaison entre mâle et femelle. Mais elle est suffisamment faible pour être négligeable.
3/ La fréquence de recombinaison varie selon les gènes étudiés Croisement à trois facteurs ou test trois point [spineless] : réduction des soies de la tête et du thorax : allèle s ; allèle s+ [Delta] : forme des veines de l’aile : allèle D dominant/ d+ (allèle sauvage); D létal à l’état homozygote [ebony] : couleur du corps : allèle e récessif / e+ (allèle sauvage). P1 Mâle s+/s+; D/d+ ; e+/e+ Femelle s/s; d+/d+ ; e/e 50% [Delta] : s+/s; D/d+ ; e+/e 50% [+] : s+/s; d+/d+ ; e+/e F1 La femelle F1[Delta] a reçu de son père les allèles (s+, D, e+), et de sa mère les allèles (s,d+,e).
Femelle F1 [Delta] : s+/s; D/d+ ; e+/e Mâle [spineless, ebony] : s/s; d+/d+ ; e/e Croisement test : [spineless, ebony] : 376 [Delta] : 380 [spineless, Delta] : 31 [ebony] : 35 [spineless] : 18 [Delta, ebony] : 21 3 couples d’allèles en ségrégation; la femelle F1 est hétérozygote pour chacun d’eux, le mâle homozygote récessif. Selon la 2ème loi de Mendel, la femelle F1 produit autant de gamètes ayant s que de gamètes ayant s+, autant de D que d+, autant de e que de e+. Selon la 3ème loi de Mendel, on devrait avoir une ségrégation indépendante des gènes, et donc 23 phénotypes possibles en proportion identiques. Or on en a que 6 phénotypes….en proportions différentes…
Femelle F1 [Delta] : s+/s; D/d+ ; e+/e Mâle [spineless, ebony] : s/s; d+/d+ ; e/e Croisement test : s/s ; d+/d+ ; e/e [spineless, ebony] : 376 s+/s ; D/d+ ; e+/e [Delta] : 380 s/s ; D/d+ ; e+/e [spineless, Delta] : 31 s+/s ; d+/d+ ; e/e [ebony] : 35 s/s ; d+/d+ ; e+/e [spineless] : 18 s+/s; D/d+ ; e/e [Delta, ebony] : 21 3 couples d’allèles en ségrégation; la femelle F1 est hétérozygote pour chacun d’eux, le mâle homozygote récessif. Selon la 2ème loi de Mendel, la femelle F1 produit autant de gamètes ayant s que de gamètes ayant s+, autant de D que d+, autant de e que de e+. Selon la 3ème loi de Mendel, on devrait avoir une ségrégation indépendante des gènes, et donc 23 phénotypes possibles en proportion identiques. Or on en a que 6 phénotypes….en proportions différentes…
Femelle F1 [Delta] : s+/s; D/d+ ; e+/e Mâle [spineless, ebony] : s/s; d+/d+ ; e/e Ségrégation des gènes s-e P s/s ; d+/d+ ; e/e [spineless, ebony] : 376 P s+/s ; D/d+ ; e+/e [Delta] : 380 s/s ; D/d+ ; e+/e [spineless, Delta] : 31 R R s+/s ; d+/d+ ; e/e [ebony] : 35 R s/s ; d+/d+ ; e+/e [spineless] : 18 R s+/s; D/d+ ; e/e [Delta, ebony] : 21 3 couples d’allèles en ségrégation; la femelle F1 est hétérozygote pour chacun d’eux, le mâle homozygote récessif. Selon la 2ème loi de Mendel, la femelle F1 produit autant de gamètes ayant s que de gamètes ayant s+, autant de D que d+, autant de e que de e+. Selon la 3ème loi de Mendel, on devrait avoir une ségrégation indépendante des gènes, et donc 23 phénotypes possibles en proportion identiques. Or on en a que 6 phénotypes….en proportions différentes… Frec (s-e) = (31 + 35 + 18 + 21)*100/861 = 12,19%
Femelle F1 [Delta] : s+/s; D/d+ ; e+/e Mâle [spineless, ebony] : s/s; d+/d+ ; e/e Ségrégation des gènes s-d s-e P P s/s; d+/d+ ; e/e [spineless, ebony] : 376 P P s+/s; D/d+ ; e+/e [Delta] : 380 s/s; D/d+ ; e+/e R [spineless, Delta] : 31 R R R s+/s; d+/d+ ; e/e [ebony] : 35 P R s/s; d+/d+ ; e+/e [spineless] : 18 P R s+/s; D/d+ ; e/e [Delta, ebony] : 21 3 couples d’allèles en ségrégation; la femelle F1 est hétérozygote pour chacun d’eux, le mâle homozygote récessif. Selon la 2ème loi de Mendel, la femelle F1 produit autant de gamètes ayant s que de gamètes ayant s+, autant de D que d+, autant de e que de e+. Selon la 3ème loi de Mendel, on devrait avoir une ségrégation indépendante des gènes, et donc 23 phénotypes possibles en proportion identiques. Or on en a que 6 phénotypes….en proportions différentes… Frec (s-d) = (31 + 35)*100/861 = 7,66%
Femelle F1 [Delta] : s+/s; D/d+ ; e+/e Mâle [spineless, ebony] : s/s; d+/d+ ; e/e Ségrégation des gènes d-e s-d s-e P P P s/s; d+/d+ ; e/e [spineless, ebony] : 376 P P P s+/s; D/d+ ; e+/e [Delta] : 380 P s/s; D/d+ ; e+/e R [spineless, Delta] : 31 R P R R s+/s; d+/d+ ; e/e [ebony] : 35 R P R s/s; d+/d+ ; e+/e [spineless] : 18 R P R s+/s; D/d+ ; e/e [Delta, ebony] : 21 3 couples d’allèles en ségrégation; la femelle F1 est hétérozygote pour chacun d’eux, le mâle homozygote récessif. Selon la 2ème loi de Mendel, la femelle F1 produit autant de gamètes ayant s que de gamètes ayant s+, autant de D que d+, autant de e que de e+. Selon la 3ème loi de Mendel, on devrait avoir une ségrégation indépendante des gènes, et donc 23 phénotypes possibles. Or on en a que 6 phénotypes…. Frec (d-e) = (18 + 21)*100/861 = 4,529%
s d e 4,529 7,66 12,19 Frec (s-d) = (31 + 35)*100/861 = 7,66% Frec (d-e) = (18 + 21)*100/861 = 4,529% Frec (s-e) = (31 + 35 + 18 + 21)*100/861 = 12,19% Carte génétique : Si les fréquences de recombinaison sont additives, cela démontre que la fréquence de CO est constante par unité de distance le long du chromosome : la fréquence de recombinaison (en % de recombinaison = cM centi Morgan) entre deux gènes peut donc être considérée comme une estimation de la distance physique (en paire de base) entre ces deux gènes
Pourquoi n’a t’on obtenu que 6 classes phénotypiques au lieu des 8 attendues? s/s; d+/d+ ; e/e [spineless, ebony] : 376 s+/s; D/d+ ; e+/e [Delta] : 380 s/s; D/d+ ; e+/e [spineless, Delta] : 31 [ebony] : 35 s+/s; d+/d+ ; e/e [spineless] : 18 s/s; d+/d+ ; e+/e [Delta, ebony] : 21 s+/s; D/d+ ; e/e
Pourquoi n’a t’on obtenu que 6 classes phénotypiques au lieu des 8 attendues? s/s; d+/d+ ; e/e [spineless, ebony] : 376 s+/s; D/d+ ; e+/e [Delta] : 380 s/s; D/d+ ; e+/e [spineless, Delta] : 31 [ebony] : 35 s+/s; d+/d+ ; e/e [spineless] : 18 s/s; d+/d+ ; e+/e [Delta, ebony] : 21 s+/s; D/d+ ; e/e s+/s; D/d+ ; e/e [spineless, Delta, ebony] : 0 s+/s; d/d+ ; e+/e [+] : 0
s d e d+ e s 4,529 7,66 d+ e s 12,19 e e d+ D s s e+ e+ D s+ s+ d+ e+ e+ D D s+ s+ s+/s; D/d+ ; e/e [spineless, Delta, ebony] : 0 s+/s; d/d+ ; e+/e [+] : 0
Pour obtenir un individu [spineless, Delta, ebony], ou un individu [+], il faut non seulement un CO entre s et D (cela se produit dans 7,66% des méïoses), mais aussi un CO entre D et e (4,529% des cas). La fréquence attendue des gamètes porteurs de ce génotype est donc de : (4,529/100) x (7,66/100) = 34,7/10000 = 0,347% S’IL S’AGIT DE 2 EVENEMENTS INDEPENDANTS Pour 861 méïoses, on s’attend à trouver (861*0,347)/100 = 3 situations de gamètes porteurs de double CO On aurait du donc avoir 1,5 (!!) mouche [spineless, Delta, ebony] et autant de phénotype sauvage • Pourquoi ne les a-t’on pas eu? • Le hasard ? • Autre chose (les 2 CO ne sont pas indépendants l’un de l’autre) ? Effectif trop faible pour répondre à cette question
[v] 580 [cv, ct] 592 [v, cv] 45 [ct] 40 [v, cv, ct] 89 [+] 94 [v, ct] 3 [cv] 5 Autre exemple !! v : œil vermillon; v+ allèle sauvage cv : absence d’une nervure dans l’aile ; cv+ allèle sauvage ct : bord des ailes soupées; ct+ allèle sauvage x Mâle [v] Femelle [ct,cv] Parents de race pure [+] Femelle F1 [+] x Mâle [ct,cv,v]
[v] 580 [cv, ct] 592 [v, cv] 45 [ct] 40 [v, cv, ct] 89 [+] 94 [v, ct] 3 [cv] 5 v : œil vermillon; v+ allèle sauvage cv : absence d’une nervure dans l’aile ; cv+ allèle sauvage ct : bord des ailes soupées; ct+ allèle sauvage x Mâle [v] Femelle [ct,cv] Parents de race pure [+] Femelle F1 [+] x Mâle [ct,cv,v] Autant de [v] que de [v+] Autant de [cv] que de [cv+] Autant de [ct] que de [ct+] • Trois gènes en ségrégation
[v] 580 [cv, ct] 592 [v, cv] 45 [ct] 40 [v, cv, ct] 89 [+] 94 [v, ct] 3 [cv] 5 v : œil vermillon; v+ allèle sauvage cv : absence d’une nervure dans l’aile ; cv+ allèle sauvage ct : bord des ailes coupées; ct+ allèle sauvage x Mâle [v] Femelle [ct,cv] Parents de race pure ct+ /ct+ ; cv+/ cv+; v/v ct /ct ; cv / cv; v+/ v+ [+] ct /ct+ ; cv / cv+; v+/v ct+ /ct ; cv+ / cv; v/v ct /ct ; cv / cv; v+/v Femelle F1 [+] ct+ /ct ; cv / cv; v/v x ct /ct ; cv+ / cv; v+/v ct /ct ; cv / cv; v/v Mâle [ct,cv,v] ct+ /ct ; cv+ / cv; v+/v ct /ct ; cv+ / cv; v/v ct+ /ct ; cv / cv; v+/v Autant de [v] que de [v+] Autant de [cv] que de [cv+] Autant de [ct] que de [ct+] • Trois gènes en ségrégation
[v] 580 [cv, ct] 592 [v, cv] 45 [ct] 40 [v, cv, ct] 89 [+] 94 [v, ct] 3 [cv] 5 Ségrégation des gènes cv-ct v-cv v-ct P P P ct+ /ct ; cv+ / cv; v/v P P P ct /ct ; cv / cv; v+/v P R R ct+ /ct ; cv / cv; v/v P R R ct /ct ; cv+ / cv; v+/v R R P ct /ct ; cv / cv; v/v R R P ct+ /ct ; cv+ / cv; v+/v ct /ct ; cv+ / cv; v/v R P R R P R ct+ /ct ; cv / cv; v+/v Frec(v-ct) = 100(89+94+3+5)/1448 = 13,2% Frec(v-cv) = 100(45+40+89+94)/1448 = 18,5% Frec(cv-ct) = 100(45+40+3+5)/1448 = 6,4% v ct cv 13,2 6,4 18,5 ou 19,6??
[v] 580 [cv, ct] 592 [v, cv] 45 [ct] 40 [v, cv, ct] 89 [+] 94 [v, ct] 3 [cv] 5 Ségrégation des gènes cv-ct v-cv v-ct P P P ct+ /ct ; cv+ / cv; v/v P P P ct /ct ; cv / cv; v+/v P R R ct+ /ct ; cv / cv; v/v P R R ct /ct ; cv+ / cv; v+/v R R P ct /ct ; cv / cv; v/v R R P ct+ /ct ; cv+ / cv; v+/v ct /ct ; cv+ / cv; v/v R P R R P R ct+ /ct ; cv / cv; v+/v Frec(v-ct) = 100(89+94+3+5)/1448 = 13,2% Frec(v-cv) = 100(45+40+89+94)/1448 = 18,5% Frec(cv-ct) = 100(45+40+3+5)/1448 = 6,4% v ct cv 13,2 6,4 18,5 ou 19,6??
[v] 580 [cv, ct] 592 [v, cv] 45 [ct] 40 [v, cv, ct] 89 [+] 94 [v, ct] 3 [cv] 5 Ségrégation des gènes cv-ct v-cv v-ct P P P ct+ /ct ; cv+ / cv; v/v P P P ct /ct ; cv / cv; v+/v P R R ct+ /ct ; cv / cv; v/v P R R ct /ct ; cv+ / cv; v+/v R R P ct /ct ; cv / cv; v/v R R P ct+ /ct ; cv+ / cv; v+/v ct /ct ; cv+ / cv; v/v R P R R P R ct+ /ct ; cv / cv; v+/v Frec(v-ct) = 100(89+94+3+5)/1448 = 13,2% Frec(v-cv) = 100(45+40+89+94 + (3 + 5)x2)/1448 = 19,61% Frec(cv-ct) = 100(45+40+3+5)/1448 = 6,4% v ct cv 13,2 cM 6,4 cM 19,6 cM
Interférence Les évènements de recombinaisons sont-ils réellement indépendants les uns des autres ? v ct cv 13,2 cM 6,4 cM 19,6 cM S’ils étaient indépendants : f(2C0) = f(1CO entre v et ct) x f(1 CO entre ct et cv) f(2C0) = 13,2% x 6,4% = 0,8448% Effectif : 1448 [v,ct] et [cv] : (0,8448x1448)/100 = 12 Or on en observe 8 Les crossing over simples sont donc favorisés par rapport aux doubles
Attendus - observés X 100 Interférence (I) = Attendus Interférence Les évènements de recombinaisons sont-ils réellement indépendants les uns des autres ? v ct cv 13,2 cM 6,4 cM 19,6 cM 8 doubles CO observés 12 doubles CO attendus entre v et cv = 1 – coef coïncidence I = 100(12-8)/12 = 33,33% Dans 1/3 des cas, le 2ème CO ne se fait pas
Signification de la liaison génétique Quelles sont les valeurs extrêmes d’une fréquence de recombinaison ? Fréquence de recombinaison de 0% ? Exemple chez la levure : [ade-, his-] x [+] 2n : [+] [ade-, his-] 492 [+] 508
Limite supérieure de la fréquence de recombinaison? Fréquence de recombinaison 50 30 0 distance génétique (cM) 10 50 60 A partir de 50cM, deux gènes sont génétiquements indépendants.
Fréquence de recombinaison 50 30 0 distance génétique (cM) 10 50 60 Dans la réalité, au delà de 30cM, il est difficile d’établir la distance entre deux gènes directement par croisement
Analyse génétique des asques Jusqu’à présent , la méiose est analysée de manière statistique • Autant de gamètes porteurs de l’allèle A que de l’allèle a.
Chez certains organismes, les produits de la méïose sont contenus dans une enveloppe
Asques produits par la levure Saccharomyces cerevisiae Asques produits par le champignon Neurospora crassa
Analyse de tétrade chez Saccharomyces cerevisiae [ura+] x [+] asques aiguille du micromanipulateur asques 1234 Milieu complet spores 1234 Milieu minimum Etapes de la dissection des asques
ura3+,his2+ ura3+,his2- ura3-,his2- ura3-,his2- ura3-,his2- ura3-,his2+ ura3+,his2+ ura3+,his2+ Analyse génétique des asques ura3-,his2+ ura3+,his2- ura3-, his2+ x ura3+, his2- ura3-/ura3+; his2-/his2+ ura3-,his2+ ura3+,his2- ura3+,his2- ura3-,his2+ T DR DP