Download

1 / 1
10 likes | 104 Views
Are There Yield Genes in Wild Soybean? Searching for Yield Potential in Lines from a Cross between Wild ( G. soja ) and Cultivated ( G. max ) Soybean. Leah Ruff 1* , Thomas Carter 2 , Dave Dickey 1 , Lilian Miranda 2 , and Julian Chaky 3

E N D
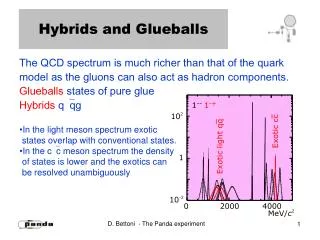
Are There Yield Genes in Wild Soybean? Searching for Yield Potential in Lines from a Cross between Wild (G. soja) and Cultivated (G. max) Soybean Leah Ruff1*, Thomas Carter2, Dave Dickey1, Lilian Miranda2, and Julian Chaky3 1Dept. of Crop Sciences, NC State University Raleigh, NC, 2USDA- ARS Raleigh, NC, 3Pioneer Hi-Bred *Leah received her bachelors at Iowa State University and is working on her master’s degree in Crop Science at NCSU Diversity in G. max x G. soja Parental Stock Introduction The breeding lines studied in this project were developed by Jacob Delheimer and Dr. Thomas Carter at North Carolina State University. They were developed from a single cross between G. max ‘N7103’ and G. soja PI 366122 in 2002 and repeated in 2003. Both breeding cycles used pedigree selection. Selection was made on their G. max–like agronomic characteristics such as, upright plant stature and branches shorter than the main stem. From the combined selection cycles, 18 recombinant inbred breeding lines (RILs) and one breeding line from Dr. Roger Boerma at the University of Georgia (F4:7 plants) were chosen on the basis of their yield and phenotypic characteristics, such as leaf shape and seed color. The three-dimensional graph in Figure 1 shows the genetic distance of the 18 inbred lines developed from the cross between G. max and G. soja. The different colors distinguish from which breeding cycle the line came. Table 1 shows that some of these lines have up to 40% G. soja alleles and yield up to 40.1 bushels/acre and within 99% of the soybean check varieties. The soybean germplasm collection available has been vastly underutilized for the improvement of soybean yield (Tanksley and McCouch 1997). More than 80% of the gene pool in the United States originates from less than a dozen soybean introductions, which implies that the genetic base is too narrow for commercial soybean [Glycine max (L.) Merr.] (Delannay et al. 1983). To guarantee future global food security and sustainable crop production, there exists a strong need for broadening the genetic base and looking for new resources to develop soybean cultivars. Exotic germplasm, such as the wild progenitor of soybean, Glycine soja, may be an excellent source of agronomic genes and genetic variability (Lee et al. 2008). Wild soybean is cross-compatible with the cultivated soybean (Singh and Hymowitz 1988). It also contains a great deal of diversity (Carter et al. 2004) and has the potential to improve soybean yield (Concibido et al. 2003). Although many papers have been published on wild soybean and on the progeny derived from it, we do not really know if it has yield genes that could increase the yield potential of cultivated varieties. The objective of this study is to determine if yield genes exist in wild soybean by examining heterosis/hybrid vigor in F2 breeding populations (Burton and Brownie 2006). These populations are derived from breeding lines which are themselves developed from a cross between Glycine max and Glycine soja. Figure 1.) Three dimensional depiction of genetic distance from the original G. max ‘N7103’and G. soja PI 366122 parents. The genotypes in blue were developed in the first breeding cycle. The green colored genotypes were developed in the second breeding cycle. G. max x G. soja Lines Used as Hybrid Parents • Examples of the G. max x G. soja F4:7 inbred lines grown in 2011. These were used as parental stock to develop hybrids • Glycine soja PI 366122 seeds • Glycine max N7103 seeds NMS4-8-146, NMS4-44-329,NMS4-112-594 NMS5-102-1-207, NMS5-70-6-129, NMS5101-2-203 Table 1.) Agronomic performance of the 19 breeding lines and the checks evaluated in three different environments. X NMS5-12-1-9,NMS5-11-1-4,NMS5-188-1-406 NMS4-169-706 • Examples of the F4:7 seeds from the advanced breeding lines G07-6012 NMS5-62-4-111 NM5-70-6-129 NMS4-44-329 NMS4-175-709 Development of Hybrids Future Analysis and Agricultural Impact None of the 19 inbred breeding lines with up to 40% G. soja pedigree exceeded the G. max conventional checks in terms of seed yield. We hypothesized that hybrids derived from them might exceed in yield, and that hybrid vigor would provide evidence of yield genes in wild soybean. During the summer of 2011, we crossed each of the 19 lines to ‘N7103’, a small-seeded maturity group VII soybean, which was also the G .max parent of the breeding lines. The F1 seed created from these crosses was sent to the winter nursery in Puerto Rico. The F1 plants of these crosses were allowed to self pollinate in Puerto Rico in the winter of 2011-2012. Tissue samples will be taken from each plant and molecular markers will be run to verify that each plant is indeed a true cross to N7103. Those that are determined not to be true crosses will be rouged. Each line’s F2 seed will be harvested separately in bulk. In the summer of 2012, the F2 seeds from the winter nursery will be planted in yield performance trials with multiple replications and in multiple locations. The locations will include Kinston and Plymouth agricultural research stations, in North Carolina, and potentially locations in Georgia and Arkansas. The flow chart below outlines this work. After the yield trials in the summer of 2012, the yield data will be compared to the yield of the G. max parent to determine if yield genes are present in Glycine soja. If the yield trials suggest hybrid vigor exists, this would be evidence to support the existence of yield genes in G. soja. Soybean breeders could use this result to direct research which incorporates new yield genes into their breeding programs and increase soybean yield. Ultimately, researchers and farmers could be working with soybeans that are higher yielding while more genetically diverse than their present soybeans. The impact of this would be to increase the soy food supply and to broaden the genetic background of soybean crops to help guarantee future crop production. Acknowledgements Thanks to Pioneer Hi-Bred and the USDA-ARS for their support and funding of this project. References Burton JW, and Brownie C. (2006) Heterosis and Inbreeding Depression in Two Soybean Single Crosses. Crop Science 46:2643-2648 Carter Jr. TE, Nelson RL, Sneller CH, Cui Z. (2004) Genetic Diversity in Soybean. In: HR Boerma and JE Specht (Eds), Soybeans: Improvement, Production, and Uses. Agronomy Monographs 3rd ed. No. 16, ASA-CSSA-SSSA, Madison, WI, USA, pp. 303–416. Delannay X, Rodgers DM, Palmer RG. (1983) Relative Genetic Contributions among Ancestral Lines to North American Soybean Cultivars. Crop Sci 23:944-949 Lee J, Yu J, Hwang Y, Blake S, So Y, Lee G, Nguyen HT, Shannon JG. (2008) Genetic Diversity of Wild Soybean (Glycine soja Sieb. and Zucc.) Accessions from South Korea and Other Countries. Crop Sci 48(2):606-16. Singh RJ, Hymowitz T. (1988) The Genomic Relationship between Glycine max (L) [Merr and G. soja Sieb. and Zucc.] as Revealed by Pachytene Chromosome Analysis. Theor Appl Genet 76:705-711 Tanksley SD, McCouch SR. (1997) Seed Banks and Molecular Maps: Unlocking Genetic Potential from the Wild. Science 277(5329):1063-6. • Hybrids (F1 plants) growing at the winter nursery in Puerto Rico on Jan. 5, 2012. These are from the crosses of advanced lines (<40% G. soja) x soybean cultivar