Download

1 / 64
640 likes | 938 Views
Molekulární biologie. 10. Mutace a opravy DNA, rekombinace ( kapitola 23, 24). MUTACE. změny genetického materiálu daného organizmu ( dědičné u potomků dané buňky ) v ětšina mutací není letální , ani se na fenotypu neprojeví . Některé mutace dokonce přinášejí organizmu výhodu .

E N D
Molekulárníbiologie 10. Mutace a opravy DNA, rekombinace (kapitola 23, 24)
MUTACE • změnygenetickéhomateriáludanéhoorganizmu (dědičné u potomkůdanébuňky) • většinamutacíneníletální, ani se nafenotypuneprojeví. Některémutacedokoncepřinášejíorganizmuvýhodu. • mutace v somatickýchbuňkáchnejsoudědičné, pouzemutace v zárodečnýchbuňkách • vetšinamutací je recesivních, efektvykompenzovándruhoualelou • Každýčlověknese v genomu 75-100 škodlivýchrecesivníchalel, z toho 8 letálních. Kdybyčlověkexistovaljako haploid, byl by dávnomrtvý! • K tomumákaždýčlověk v genomuasi 20.000 změn, kteréjsou ale neškodné • Nulovámutace – úplněchybíproduktdaného genu, loss of function • Tichámutace – nemávlivnafunkcidaného genu, bezfenotypu, např. vetřetípozicikodonu, v intronech, v intergenové DNA • Substitucebáze • Inserce • Delece • Inverse • Duplikace • Translokace uvnitř genu nebo v regulačníchsekvencích
Substitucebáze bodovámutace
Missensemutace– změníkódovanouaminokyselinu v proteinu radikálnímutace (nahrazeníaminokyselinou s velmirozdílnýmivlastnostmi, vlivnastrukturu a tímfunkciproteinu) neutrálnímutace (nahrazeníaminokyselinou s podobnýmivlastnostmi) Aminokyselinykonzervovanéu rozdínýchorganizmůbudou dost možná v aktivnímmístěproteinu a jejichmutacebudemítzávažnějšídůsledkynežmutaceostatníchaminokyselin.
Kondicionálnímutace – efektzáležínapodmínkách, vekterýchorganizmusžije např. teplotněsensitivnímutace – projeví se jenpřiurčitýchteplotách Mutace v melaninu, která ho děláaktivnějšímvchladnějšíchčástechtěla.
Nonsensemutace – zavedenípředčasného stop kodonu v mRNA např. UCG UAG ser stop Prokaryota – protein je syntetizovánaž do předčasného stop kodonu, pakuvolněn, ale protoževětšinounenísprávněsložený, je degradován Eukaryota – nonsense mediated mRNA decay, zničí se rovnou mRNA Nonsense mutace = většinouefektjakonulovámutace
Delece Efektvětšídelecemůžebuďinaktivovatgenyneboijejichexpresizvýšit (delecevazebnéhomísta pro represor) Delecejednéčiněkolikabazí – frameshiftvečtecímrámcinebovlivna splicing, vlivnavazbutranskripčníchfaktorůatd.
Frameshift -1 Frameshift -2 Frameshift -3
Inzerce Inzercemobilníhogenetickéhoelementu (transposony, retrotransposony), virů, nebochemickýmimutagenyčichybou DNA polymerázy zastaveníexprese zvýšeníexprese
Inverze Translokace Duplikace
Spontánnímutace chyby v DNA replikaci, spontánníchemickézměnyna DNA (s nízkoufrekvencí, se zvyšující se teplotoufrekvenceroste) Indukovanémutace Mutageny: chemickélátky, radiace, teplo Chemickémutageny: • EMS (ethyl methane sulphonate) • alkylačníčinidlo, přidáváethylovéskupinybazím • pro in vivo mutageneziživýchbuněk • Dusitany(NO2-) • přeměnaaminoskupinnahydroxyskupiny • mutace5-methyl cytosinunathymin • pro in vitro mutageneziplazmidů
Analogy bází • 5-bromouracyl • inkorporuje se do DNA běhemreplikacejakothymin • existujevedvou flip-flop stavech, z nichžjeden se páruje s A a jeden s G • problémpřinásledné DNA replikaci, kdysloužíjakotemplát
Interkalačníčinidla • acridine orange, acriflavin, ethidiumbromid • vmezeří se mezidvěbáze DNA • DNA polymerázaběhemreplikacepakrozeznátutolátkujakoskutečnoubázi a vloží extra nukleotid do DNA, frameshift… Teratogen – látkazpůsobujícívývojovévadyembryí (buďjako mutagen nebojinýmimechanizmy)
Mutacezpůsobenéradiací: • elektromagentickévlnění s velmivysokoufrekvencí(UV) • UV světlomávlnovoudélku 100-400nm, bázeabsorbujínejvícepřivlnovédélce 254nm, tvorbathyminovýchdimérů, problémpřireplikaci • většina UV zesluncezachycenaozonovouvrstvou, pokudnenípoškozená… Thyminovédiméry thymin thyminový dimer • ionizačnízáření(X-ray, g-ray) • přistyku s vodoučijinýmilátkamiindukujítvorbuiontů a volnýchradikálů (hlavně OH.) = nepřímépoškození DNA • taképřímépoškození DNA, přímoureakcízáření s DNA (hlavně ds zlomy)
SPONTÁNNÍ MUTACE Mutacevznikléchybami DNA polymerázy • proofreading aktivita DNA polymerázynenídokonalá, s maloufrekvencízůstávajíbehěmreplikace v DNA chyby • uE.colibehěmreplikace leading stranduzařadípolymerázaprůměně 1 špatnoubázi z 10 milionůzreplikovaných, u lagging strandu 20x více (protože DNA PolImámáhorší proofreading aktivitu) • navícmůžedocházeti k ‘uklouznutí’ polymerázy (polymerase slipping): zavedenídelecíneboinzercí
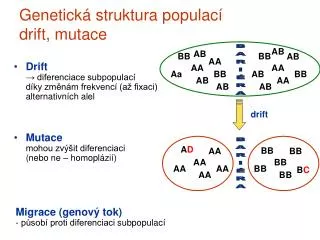
Mutacekvůlipárovánípodobnýchúseků DNA a následnérekombinaci přímérepeticena DNA rekombinace(mezidvěmatotožnýminebovelmipodobnýmisekvencemi) DELECE a DUPLIKACE DELECE
obrácenérepeticena DNA INVERZE
Mutacekvůlitautomerizacibazí • každábázeexistujejakoketo a enol forma – tautomery • vevzájemnérovnováze, ale značněpřevažujíketoformy • pokudpřireplikacibázezrovna v enolformě, budezařazena v nesprávnémpárování
Mutacekvůlispontánníchemickéinstabilitěbazí A, G a C mohoupomaluspontánněztrácetsvéaminoskupiny = DEAMINACE 100 bazíza den v každéeukaryotníbuňce! záměnaCzaT Cytosin je horkýmmístemmutací v DNA ! deaminace G a A mnohemvzácnější
Mutacekvůlispontánníchemickéinstabilitěbazí A a G se mohouspontánněhydrolyzovat od DNA kostry = DEPURINACE 5000 bazíza den v každéeukaryotníbuňce!
Poškozeníoxidativnímstresem(volnýmiradikály, např. běhembuněčnéhometabolismu) formace8-hydroxyguaninu párování s A záměna G/C za A/T Poškozeníneenzymatickoumetylací S maloufrekvencímůže donor metylačníchskupin (S-adenosylmethionin) spontánněmetylovatbáze adenin 3-methyladenin
OPRAVY DNA Prokaryota
Proofreading aktivitaDNA polymeráz se snažíeliminovatšpatnépárováníběhemreplikace Mismatch v párováníbazí, jejichchemickémodifikacenebovýskytanalogůbazívede k distorci DNA Mismatch repair system a excision repair system detekujítytozměnystruktury DNA spíšenežspecifickéchemickézměny Jakreparačnísystémypoznají, kterázedvounesprávněspárovanýchbazí je ta správná?
Jakreparačnísystémypoznají, kterázedvounesprávněspárovanýchbazí je ta správná? Je třebaumětrozeznatpřireplikaciparentální DNA vlákno od nověreplikovaného Bakteriální DNA metylázymetylujíparentálnívlákno DNA dam (DNA adenine methylase) - GATC dcm(DNA cytosine methylase) – CCAGG nebo CCTGG 6-methyladenin a 5-methylcytosin se párujísprávně, neindukujíreparačníodpověd’
několikminutporeplikaci je nová DNA šroubovicehemimetylovaná nastupujíreparačníenzymy a hledajíchyby v nemetylovanémvlákně dam a dcmmetylázymetylujíinovévlákno
DNA mismatch repair system (MMR system) rozpoznáníchybnébázepomocíMutL , MutS a MutHproteinů zavedenízlomu v novém DNA vlákně odstraněníúseku DNA kolemchybnéhopárování dosyntetozovánísprávnébáze DNA polymerázou III (nebod u eukaryot)
Nucleotide excision repair system (NER system) • nejčastějšísystémopravpoškozené DNA • rozpoznávázměnystruktury DNA, ale méněsenzitivnínež MMR (změnystrukturymusejíbýtvícenápadné) • předevšímopravy DNA poškozenéUV = thyminovédimery, kroslinkovanébáze rozpoznáníchybnébázepomocíuvrA,B,Cproteinů zavedenídvouzlomůokolo T=T odstraněníúseku DNA kolemchybnéhopárování a dosyntetozovánísprávnébáze DNA polymerázouI
Base excision repair system (BER system) • rozeznáváspecifickéchemickézměny v DNA, která se neprojevínazměněstruktury DNA • opravujebáze, které se normálně v DNA nevyskytují (nenípochyb o tom, žejsoušpatné) Deaminaceadeninunahypoxantin, guaninunaxanthin, cytosinunauracil: DNA glykosylázaodštěpíchybnoubázi, vzniknemístobezbáze (AP místo) AP endonukleázaštěpíkostru DNA DNA polymeráza I dostavímezeru
Oxidaceguaninuna 8-oxoguanin: MutTfosfatázaodstraňujefosfátovéskupiny z oxoGTPnukleotidů, aby se nemohlyinkorporovat do DNA MutMglycosylázaodstraňujeoxoG z DNA (pokud se párujesprávně s C) MutYglycosylázaodstraňujeA z DNA pokud se páruje s oxoG Vzniklé AP místodostavěné DNA polymerázou I
Very short patch repair system Thyminvzniklýspontánnídeaminací 5-methyl cytosinu (ne běhemreplikace DNA) je horkýmmístemmutací (změna CG naTA) • uE.coli je 5-methylcytosin hlavně v místechmetylovaných dam a dcm • pokud se T vyskytne v sekvencíchrozeznávaných dam a dcmmetylázami (CCAGG a CCTGG), je odstraněnVsrendonukleázou Sebevražednédemetylázy Metylovéskupinynakyslíku u O6-methylguaninu a O4-methylthyminu odstraněnysebevražednýmienzymy, kterémetylovouskupinupřenesounasebe. Báze je tímrovnouopravena, ale enzymlzepoužítpouzejednou. Ostatnímetylovanébázeodstraněny DNA glycosylázami
Photoreaktivačnísystém opravathyminovýchdimerů(opravytaképomocí NER systému, Uvrproteiny) Přímáoprava, rozštěpenídimeru, bezpoužitípolymerázy Fotolyáza– absorbujeviditelnésvětlo o vlnovédélce 350-500nm (modré) a jehoenergiipoužívározštěpeníthyminovéhodimeru = fotoreaktivace
Opravypomocírekombinace Ne všechnychyby DNA se podaříodstranitpředprůběhemreplikace. Pokudmutacebránípostupu DNA polymerázy (napříkladpřítomnostthyminovýchdimerů), polymerázaodpadne a začnesyntézuopět o kousekdál Vzniknejednořetězcovámezera v replikovanémchromozomu RecA Rekombinacemezi ‘zdravým’ vláknemnadruhémchromozomu a vláknem s mezerou Mezeradostavenapodlezdravéhovlákna, thyminový dimer siceneopravený, ale obachromozomyzreplikoványbezmezer
SOS reparačnísystém Pokud je DNA poškozenánamnohamístech a běhemreplikacevzniká hodně jednovláknovýchoblastí, SOS systémumožníprůběhreplikacedosyntetizovánímtěchtoúseků, ikdyž to pravděpodobnězavedemnohomutací RecA se aktivujevazbounassDNA lexA dimer blokuje expresi SOS genů RecArozštípeLexArepresor lexA je zničen, spustí se exprese SOS genů např. DNA polymeráza V, nemá proofreading activitu, takžemůžreplikovatithyminovédimery a místabezbází (AP místa) Pouze u bakterií, pro Eukaryotapřílišnebezpečnéreplikovatbuňky s mnohamutacemi (rakovina), radějiapoptozaneboinhibicedělení
Opravaspřažená s transkripcí (transcription coupled repair) Pokud je DNA hodně poškozená, můžebránitprůběhutranskripce, RNA polymeráza se zastaví, začneopravatemplátovéhovlákna Bakterie: TRCF protein rozpoznávázastavenoupolymerázu (transcription repair coupling factor) Eukaryota: TFIIHrozvolňuje DNA běhemtranskripce, pokud’narazínapoškozenou DNA, navážeproteiny excision repair systému (NER)
Opravy u Eukaryot Systémypodobnéjako u prokaryot, ale méněprozkoumané Mutace v reparačníchgenechjsoudoprovázenévyššívýskytemrakoviny: hMSH2 (human MutS homologue 2) – mismatch repair system, vznik malýchdelecí a inzercí BRCA1 (breast cancer A1) – zlomydsDNA, transkripčníreparace Xerodermapigmentosum mutace v genech pro excision repair system, vysokásenzitivitakůže k UV záření
Problémy s opravami DNA vedoukezvýšenénáchylnosti k rakovině a k předčasnémustárnutí Wernerůvsyndrom PředčasnéstárnutízpůsobenémutacíveWnr genu – specifickéhelikázypoužívanépřiopravách DNA
DNA damage checkpoints Přikaždémdělení se napřechodech G1/S a G2/M a také v S kontrolujeintaktnost DNA ATMkináza – detekujedvojřetězcovézlomy ATRkináza – detekujezastavenéreplikačnívidličky Fosofrylujícelouřaduproteinůvedoucí k zastaveníbuněčnéhocykluneboapoptosenapř. p53
Opravydvojřetězcovýchzlomů Non homologous end joining ZlomydsDNAionizačnímzářením, chemickynebopovyštěpenítranspozonů eukaryota VazbaKuproteinůnakoncedsDNA VazbaDNA-PK (DNA protein kinasy) Fosforylace XRCC4, díkytomunavázáníligázy a spojení DNA Můžespojitomylemi DNA, která k soběnepatří, chromozomálnítranslokace dsDNAzlomymožnoopravittéžhomolognírekombinací(vizdále)
Horkámístamutací Ne všechnamísta v genomujsoustejněnáchylná k mutacím, některámístamajífrekvencidalekovyšší. Vetšinoumístavýskytu5-methylcytosinu, kterýobčasspontánnědeaminujenathymin a páruje se pak s A místo s G.
Reverzefenotypu Fenotypvzniklýurčitoumutací je vrácen do normálníhostavu Šance, že by danýnukleotidspontánnězmutovalzpětnapůvodnínukleotid je velmimalá Mnohemčastěji je reverzefenotypuzpůsobenajinoumutací, kterávyrušíúčinekprvní = supresorovámutace(aťužvestejném genu nebo v jiném)
SupresorovétRNA • nabitétRNA, které ale majízmutovanáantikodon, takžerozeznává STOP kodon(např. tRNA pro glycin, kdeantikodon GAC zmutovalna AUC) • Mohoučástečněobnovitexpresigenů s předčasným stop kodonem, běžné u prokaryot a kvasinek • PouzepokudexistujevícetRNA pro tutéžaminokyselinou, jinak by mutacekodonubylaletální Důsledkem je prodlouženíinormálníchproteinů, cožmůžebýtproblém.
Rekombinace Výměnagenetickéhomateriálumezichromozomynebonebojinýmimolekulami DNA U eukaryotpřimeioze, přiopravách DNA, u bakteriípřikonjugaciplazmidu crossing over – zlom v DNA a jejíspojení s jinoumolekulou (zdedvojitýcrossing over)
Homolognírekombinace - mezidvěmavelmipodobnýmiskevencemi Nehomologní (místněspecifická) rekombinace – mezijinaknepodobnýmisekvencemi, ale iniciovánonakrátkémúsekuhomologierozpoznávanémspecifickýmiproteiny
Holliday junction Crossing over dvoujednovláknovýchřetězců DNA a jehoposunpodélchromozomu Následujedalšíštěpení a separacevláken, může a nemusívést k výměněcelých ramen nachromozomu:
Holliday junction se můžeotáčet do dvouizomerníchstavů Patch recombinant Vymění se pouzečástjednohovlákna resolváza (rekombináza) – štěpí a liguje DNA v Holliday junction True recombinant Vymění se celýzbytekchromozomu
Crossingover / video http://booksite.academicpress.com/Clark/molecular2/anim24_crossoverformation.php
Jak se homolognísekvencenajdou a spárují ? Bakterie: zlom v dsDNA a invazejednořetězcovéhovlákna 1. RecBCDkomplexnasedánazlomydsDNA 2. Postupujepo DNA, dokudnenarazína Chi sekvenci(GCTGGTGG, časté) 3. RecCodštěpíjeden z řetězců DNA 4. RecBCpostupněodhalujessDNA s volným 3’koncem, ta se pokrývá RecAproteiny:
Zlomy v dsDNA = většinou ne zlomy v genomické DNA, ale v cizorodéDNA vniklédo buňkypřitransformaci, transdukciviry, konjugaci ssDNAvlákno s volným 3’koncem se pokrýváRecAproteiny … a atakujedsDNAhomologní DNA molekuly Dočasnévytvořenítriple helixu VytěsněnéssDNAvláknose pakspojí s ssDNAdruhémolekuly