Download
1 / 1
10 likes | 166 Views
Seed 4 fibroblast cell lines: Two human, two mice. METHODS. IMR90. Skin sh Ets-1 1. WI38. Skin. Induce senescence via IR and doxo (2x each line/treatment). Amplify human and mouse oe Ets-1 plasmid DNA. Day 0:. Collect ctr cell RNA (2x each line).

E N D
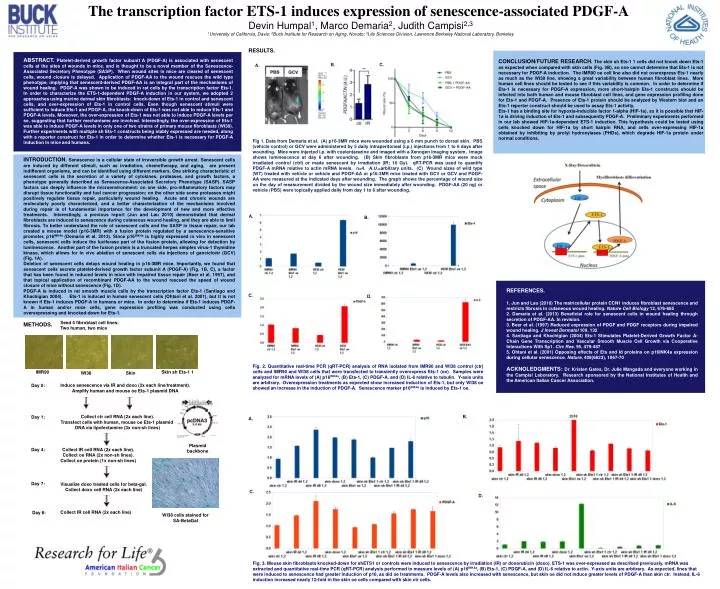
Seed 4 fibroblast cell lines: Two human, two mice METHODS. IMR90 Skin sh Ets-1 1 WI38 Skin Induce senescence via IR and doxo (2x each line/treatment). Amplify human and mouse oe Ets-1 plasmid DNA Day 0: Collect ctr cell RNA (2x each line). Transfect cells with human, mouse oe Ets-1 plasmid DNA via lipofectamine (3x non-sh lines) Day 1: Plasmid backbone Day 4: Collect IR cell RNA (2x each line). Collect oe RNA (2x non-sh lines). Collect oe protein (1x non-sh lines) Day 7: Visualize doxo treated cells for beta-gal. Collect doxo cell RNA (2x each line) Collect IR cell RNA (2x each line) Day 8: WI38 cells stained for SA-BetaGal The transcription factor ETS-1 induces expression of senescence-associated PDGF-A Devin Humpal1, Marco Demaria2, Judith Campisi2,3 1University of California, Davis; 2Buck Institute for Research on Aging, Novato; 3Life Sciences Division, Lawrence Berkeley National Laboratory, Berkeley RESULTS. ABSTRACT. Platelet-derived growth factor subunit A (PDGF-A) is associated with senescent cells at the sites of wounds in mice, and is thought to be a novel member of the Senescence-Associated Secretory Phenotype (SASP). When wound sites in mice are cleared of senescent cells, wound closure is delayed. Application of PDGF-AA to the wound rescues the wild type phenotype, implying that senescent-derived PDGF-AA is an integral part of the mechanisms of wound healing. PDGF-A was shown to be induced in rat cells by the transcription factor Ets-1. In order to characterize the ETS-1-dependent PDGF-A induction in our system, we adopted 2 approaches using murine dermal skin fibroblasts: knock-down of Ets-1 in control and senescent cells, and over-expression of Ets-1 in control cells. Even though senescent stimuli were sufficient to induce Ets-1 and PDGF-A, the knock-down of Ets-1 was not able to reduce Ets-1 and PDGF-A levels. Moreover, the over-expression of Ets-1 was not able to induce PDGF-A levels per se, suggesting that further mechanisms are involved. Interestingly, the over-expression of Ets-1 was able to induce PDGF-A levels in only one of two strains of primary mouse fibroblasts (WI38). Further experiments with multiple sh Ets-1 constructs being stably expressed are needed, along with a reporter construct for Ets-1 in order to determine whether Ets-1 is necessary for PDGF-A induction in mice and humans. CONCLUSION/FUTURE RESEARCH. The skin sh Ets-1 1 cells did not knock down Ets-1 as expected when compared with skin cells (Fig. 3B), so one cannot determine that Ets-1 is not necessary for PDGF-A induction. The IMR90 oe cell line also did not overexpress Ets-1 nearly as much as the WI38 line, showing a great variability between human fibroblast lines. More human cell lines should be tested to see if this variability is common. In order to determine if Ets-1 is necessary for PDGF-A expression, more short-hairpin Ets-1 constructs should be infected into both human and mouse fibroblast cell lines, and gene expression profiling done for Ets-1 and PDGF-A. Presence of Ets-1 protein should be analyzed by Western blot and an Ets-1 reporter construct should be used to assay Ets-1 activity. Ets-1 has a binding site for hypoxia-inducible factor 1-alpha (HIF-1a), so it is possible that HIF-1a is driving induction of Ets-1 and subsequently PDGF-A. Preliminary experiments performed in our lab showed HIF-1a-dependent ETS-1 induction. This hypothesis could be tested using cells knocked down for HIF-1a by short hairpin RNA, and cells over-expressing HIF-1a obtained by inhibiting by prolyl hydroxylases (PHDs), which degrade HIF-1a protein under normal conditions. B. C. A. Fig 1. Data from Demaria et al. (A) p16-3MR mice were wounded using a 6 mm punch to dorsal skin. PBS (vehicle control) or GCV were administered by 5 daily intraperitoneal (i.p.) injections from 1 to 6 days after wounding. Mice were injected i.p. with coelentarazine and imaged with a Xenogen imaging system. Image shows luminescence at day 6 after wounding. (B) Skin fibroblasts from p16-3MR mice were mock irradiated control (ctrl) or made senescent by irradiation (IR; 10 Gy). qRT-PCR was used to quantify PDGF-A mRNA relative to actin mRNA levels. n=4. A.U.=arbitrary units. (C) Wound sizes of wild type (WT) treated with vehicle or vehicle and PDGF-AA or p16-3MR mice treated with GCV or GCV and PDGF-AA were measured at the indicated days after wounding. The graph shows the percentage of wound size on the day of measurement divided by the wound size immediately after wounding. PDGF-AA (20 ng) or vehicle (PBS) were topically applied daily from day 1 to 6 after wounding. INTRODUCTION. Senescence is a cellular state of irreversible growth arrest. Senescent cells are induced by different stimuli, such as irradiation, chemotherapy, and aging, are present indifferent organisms, and can be identified using different markers. One striking characteristic of senescent cells is the secretion of a variety of cytokines, proteases, and growth factors, a phenotype generally described as Senescence-Associated Secretory Phenotype (SASP). SASP factors can deeply influence the microenvironment: on one side, pro-inflammatory factors may disrupt tissue functionality and fuel cancer progression; on the other side some proteases might positively regulate tissue repair, particularly wound healing. Acute and chronic wounds are molecularly poorly characterized, and a better characterization of the mechanisms involved during repair is of fundamental importance for the development of new and more effective treatments. Interestingly, a previous report (Jun and Lau 2010) demonstrated that dermal fibroblasts are induced to senescence during cutaneous wound healing, and they are able to limit fibrosis. To better understand the role of senescent cells and the SASP in tissue repair, our lab created a mouse model (p16-3MR) with a fusion protein regulated by a senescence-sensitive promoter, p16INK4a (Demaria et al. 2013). Since p16INK4a is highly expressed in vivo in senescent cells, senescent cells induce the luciferase part of the fusion protein, allowing for detection by luminescence. Another part of the fusion protein is a truncated herpes simplex virus-1 thymidine kinase, which allows for in vivo ablation of senescent cells via injections of ganciclovir (GCV) (Fig. 1A). Deletion of senescent cells delays wound healing in p16-3MR mice. Importantly, we found that senescent cells secrete platelet-derived growth factor subunit A (PDGF-A) (Fig. 1B, C), a factor that has been found in reduced levels in mice with impaired tissue repair (Beer et al. 1997), and that topical application of recombinant PDGF-AA to the wound rescued the speed of wound closure of mice without senescence (Fig. 1D). PDGF-A is induced in rat smooth muscle cells by the transcription factor Ets-1 (Santiago and Khachigian 2004). Ets-1 is induced in human senescent cells (Ohtani et al. 2001), but it is not known if Ets-1 induces PDGF-A in humans or mice. In order to determine if Ets-1 induces PDGF-A in human and/or mice cells, gene expression profiling was conducted using cells overexpressing and knocked-down for Ets-1. A. B. REFERENCES. 1. Jun and Lau (2010) The matricellular protein CCN1 induces fibroblast senescence and restricts fibrosis in cutaneous wound healing. Nature Cell Biology 12, 676-685 2. Demaria et al. (2013) Beneficial role for senescent cells in wound healing through secretion of PDGF-AA. In revision. 3. Beer et al. (1997) Reduced expression of PDGF and PDGF receptors during impaired wound healing. J Invest Dermatol 109, 132 4. Santiago and Khachigian (2004) Ets-1 Stimulates Platelet-Derived Growth Factor A-Chain Gene Transcription and Vascular Smooth Muscle Cell Growth via Cooperative Interactions With Sp1. Circ Res. 95, 479-487 5. Ohtani et al. (2001) Opposing effects of Ets and Id proteins on p16INK4a expression during cellular senescence. Nature, 409(6823), 1067-70 C. D. Fig. 2. Quantitative real-time PCR (qRT-PCR) analysis of RNA isolated from IMR90 and WI38 control (ctr) cells and IMR90 and WI38 cells that were transfected to transiently overexpress Ets-1 (oe). Samples were analyzed for mRNA levels of (A) p16INK4a, (B) Ets-1, (C) PDGF-A, and (D) IL-6 relative to tubulin. Y-axis units are arbitrary. Overexpression treatments as expected show increased induction of Ets-1, but only WI38 oe showed an increase in the induction of PDGF-A. Senescence marker p16INK4a is induced by Ets-1 oe. ACKNOLEDGMENTS:Dr. Kristen Gates, Dr. Julie Mangada and everyone working in the Campisi Laboratory. Research sponsored by the National Institutes of Health and the American Italian Cancer Association. B. A. C. D. Fig. 3. Mouse skin fibroblasts knocked-down for shETS1 or controls were induced to senescence by irradiation (IR) or doxorubicin (doxo). ETS-1 was over-expressed as described previously. mRNA was extracted and quantitative real-time PCR (qRT-PCR) analysis performed to measure levels of (A) p16INK4a, (B) Ets-1, (C) PDGF-A, and (D) IL-6 relative to actin. Y-axis units are arbitrary. As expected, lines that were induced to senescence had greater induction of p16, as did oe treatments. PDGF-A levels also increased with senescence, but skin oe did not induce greater levels of PDGF-A than skin ctr. Instead, IL-6 induction increased nearly 12-fold in the skin oe cells compared with skin ctr cells.